Популяционная генетика послеледниковой Евразии.
2 поста
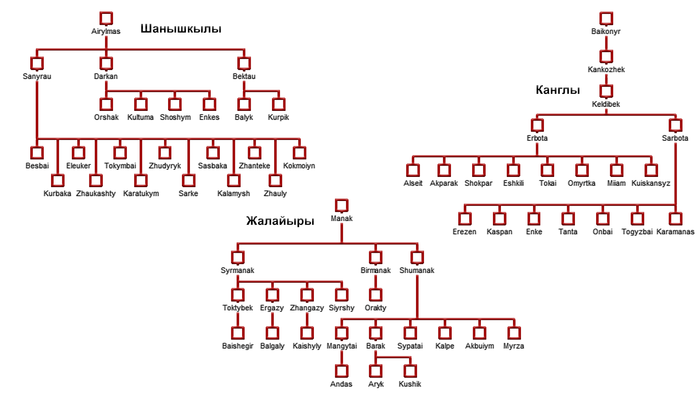
Казахи имеют одну из самых крупных родоплеменных структур в Евразийской степи. Она состоит из трёх жузов – Старший, Средний и Младший.
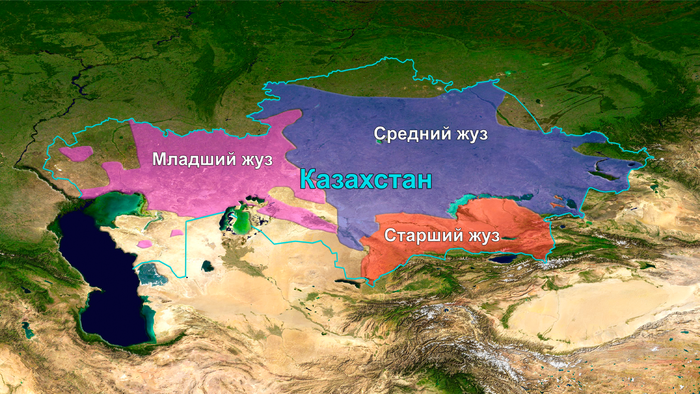
Двенадцать родов Старшего жуза в основном проживают в Южном Казахстане. Согласно традиционной генеалогии казахов, известной также как шежире, 9 из 12 родов имеют общего предка, известного как Майкы-бий.
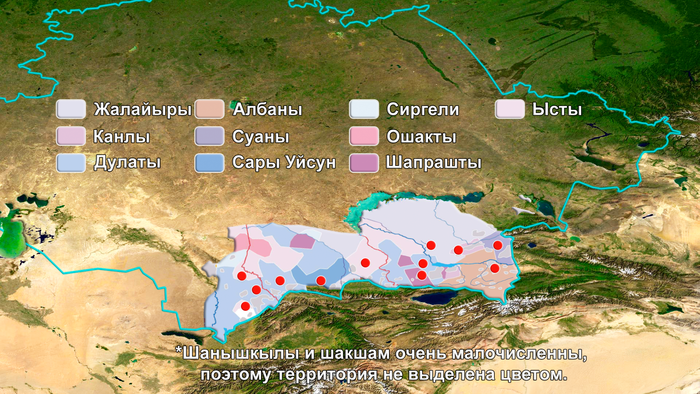
Исторические источники упоминают, что он возглавлял западную часть Золотой Орды при внуке Чингисхана, Батые. Эти девять родов вместе образуют племя уйсунов.
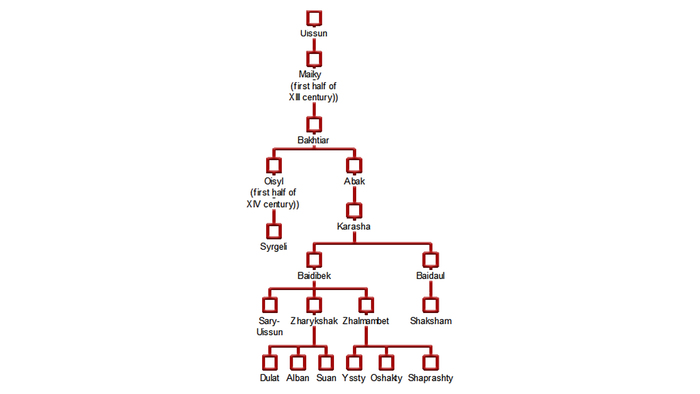
Три оставшихся рода (жалайыры, канлы и шанышкылы), имеют своих собственных предков и считаются генеалогически не связанными друг с другом и с родоплеменным объединением уйсунов.
Существует три основные гипотезы происхождения племени уйсунов. Первая - это происхождение от ранних усуни, племени индоиранского или тохарского происхождения, которое проживало в бассейне реки Тарим. Этот район примыкает к месту нынешней резиденции уйсунов.
Две другие версии предполагают более отдалённую миграцию из Монголии. Поэтому в новой работе исследователи решили проверить какая из трёх версий наиболее вероятна с точки зрения генетики.
Отцовские линии южно-казахских родов
У 490 генотипированных из 11 кланов Южного Казахстана, включая 8 из племени уйсунов, было определено 27 гаплогрупп Y-хромосомы. Что уже интересно...
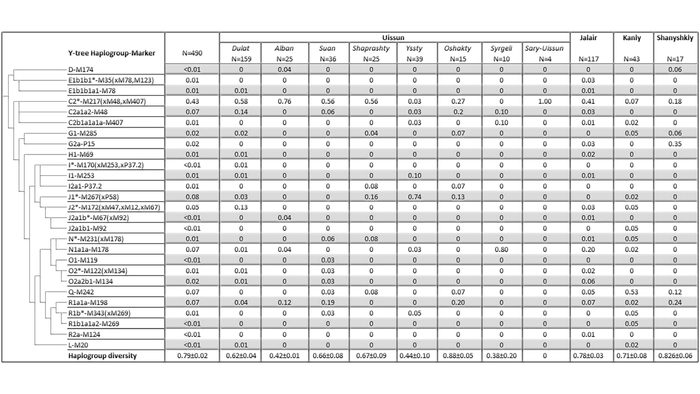
Более половины генофонда (51%) южных казахов из Старшего жуза характеризуется тремя кладами гаплогруппы Y-хромосомы С2. А 34% генофонда состоит из гаплогрупп J-M172 (13%), а также по 7% N1a1a-M178, Q-M242, и R1a1a-M198. Для большинства казахов Старшего жуза основной гаплогруппой является С2*-М217 (хМ48, xM407). Среди восьми родов племени уйсунов С2-М217 наиболее часто встречается в 6 из них (дулат, албан, суан, шапрашты, ошакты и сары уйсын). Эта гаплогруппа также характерна и для племени жалайыр. А для двух оставшихся племён уйсунов основными гаплогруппами являются N1a1a-M178 (80% у племени сиргели) и J1*-M267(xP58) (74% племени ысты). А для неуйсунских племён шанышкылов и канглов, характерны гаплогруппы Q-M242 (53% у канглов) и G2-P15 (35% для шанышкылов). Примечательно, что для последних, результаты новой работы оказались довольно противоречивы, потому как у шанышкылов из предыдущих работ была отмечена высокая частота гаплогруппы С2, в отличие от 18% в новой работе.
Структурирование отцовского генофонда уйсунов
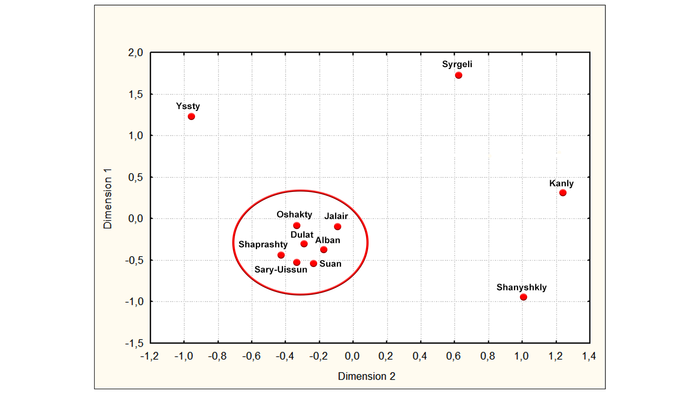
Генетические портреты трёх уйсунских родов (ысты, сиргели и ошакты) не вписываются в генеалогию, предполагающую происхождение всех девяти уйсунских родов от общего предка по имени Майкы-бий.
Генетически отличительное происхождение также подтверждается анализом молекулярной дисперсии (AMOVA). В итоге получилось объединить только семь родов в один кластер с общим происхождением: сары-уйсын, дулат, албан, суан, шапрашты, ошакты и жалайыры, при этом последний не относится к уйсунам.
Филогенетический анализ гаплогруппы С2*-F3796
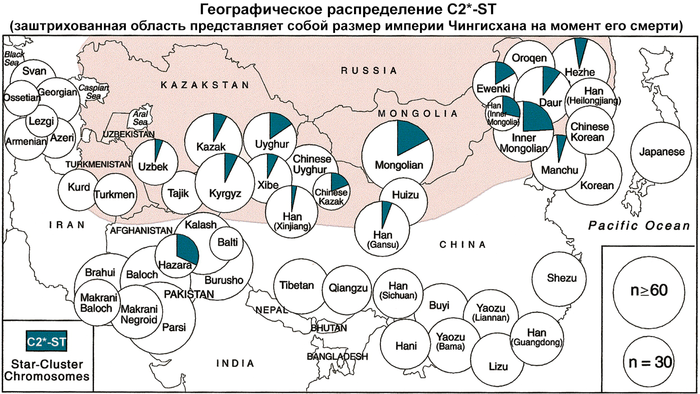
Гаплогруппа С2*-ST чётко различается в пределах M217(xM48, M407) по гаплотипам коротких тандемных повторов. Эта линия быстро распространилась по степям Евразии во время завоеваний Монгольской империи.
Этот гаплотип предположительно, был связан с гаплотипом Чингисхана или его родственников.
Наиболее высокие частоты С2*-F3796 были обнаружены у казахов из племени кереев Среднего жуза (77%), а также баргутов (46%), хазарейцев (38%), узбеков из Афганистана (35%) и монголов (35%). Наибольшее разнообразие гаплотипов в С2*-F3796 характерно для монголов и узбеков, а наименьшее для казахов.
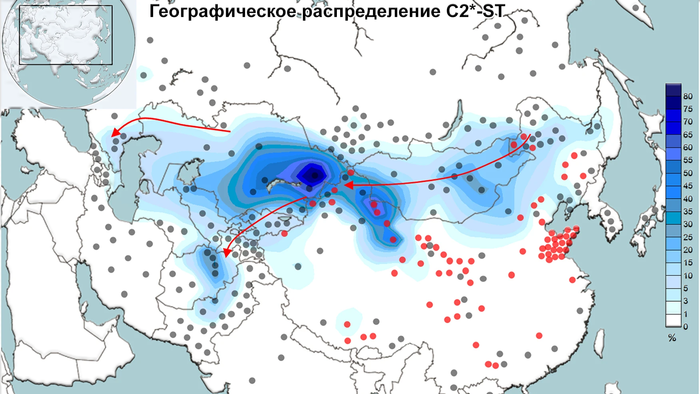
Филогенетическая сеть гаплотипов внутри гаплогруппы С2*-F3796 была построена с использованием 15 локусов коротких тандемных повторов Y-хромосомы по данным 743 человек из 25 популяций Евразии. В Сети отчётливо видны пять различных кластера.
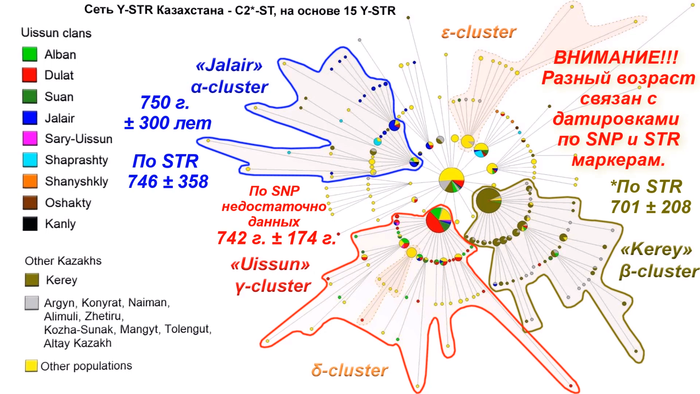
Большинство образцов из Старшего жуза Южного Казахстана были включены в два вновь выявленных подкластера: α и γ. И только несколько образцов вошли в β-кластер, датируемый 701 г. ± 208 лет. Этот кластер ранее был отнесён к племени керей Среднего жуза. α-кластер, датированный 746 г. ± 358 лет состоял в основном из членов неуйсунского рода жалайыр. А γ-кластер, который назван уйсунским кластером и датирован 742 г. ± 174 г., включает 4 уйсунских рода, а именно: дулат, албан, суан и сары уйсын. Среди двух хазарейских субкластеров δ-кластер происходит от уйсунского γ-кластера, в то время как ε-кластер происходит от общего основателя С2*-F3796 и помимо хазарейцев включает несколько узбекских гаплотипов.
Время до самого последнего общего предка
Время до самого последнего общего предка гаплогруппы С2*-F3796 по маркерам однонуклеотидного полиморфизма и коротких тандемных повторов, отличается и варьируется от 1544 лет ± 424 г., до ~ 2600 лет назад. Возраст α-кластера, составил около 750 г. ± 300 лет, возраст β-кластера ~ 650 лет ± 332 г., а γ-кластера 742 г. ± 174 г. Близкое совпадение возрастов всех трёх кластеров свидетельствует о быстром росте численности казахских родов в XIII-XIV вв. Это также прекрасно совпадает с периодом расширения Монгольской империи. Примечательно, что возраст γ-кластера “уйсун” совпадает со временем жизни предполагаемого предка Майкы-бия XIII век.
Чьими потомками являются представители Южного Казахстана?
Установлено, что субклад С2-F3796 гаплогруппы С2*-ST является наиболее распространённым в популяции Южного Казахстана. В этом смысле С2*-ST является ключом к расшифровке прямого отцовского предка родов Старшего жуза.
Более того, согласно историческим исследованиям, время жизни легендарного предка уйсунов Майкы-бия совпадает с уйсунским γ-кластером. Самому древнему образцу этой линии (подкласс С2-Y4580*) 700 лет, и он принадлежит монгольскому буддийскому погребению Золотой Орды в Центральном Казахстане из Карасуырского могильника. И он тесно связан с уйсунской гаплогруппой С2*-ST. А вот единственный изученный на сегодняшний день образец усуней из могильника Тургень-II из Семиречья, относится к гаплогруппе R1a1a-Z93(xZ94) (подкласс R1a1a-Y41571). Другие древние экземпляры из Таримского бассейна, где жили усуни, также принадлежали к гаплогруппе R1a1. При этом, все ранее изученные казахские образцы принадлежали к другой ветви R1a, а именно R1a1a-Z94 (субклад Z2125).
В целом, R1a редко встречается среди уйсунов (только 6%), поэтому их отцовские линии, скорее всего, произошли от ранних монгольских популяций, а не от представителей племени усуни.
Согласно "Сокровенному сказанию монголов", ранние монголы делились на нирун- и дарлекин-монголов. Но кто из них является предком уйсунов? Единственный генетически изученный род-преемник дарлекин-монголов – это конграты, у которых гаплогруппа С2-М407 присутствует с высокой частотой (86%), в отличие от уйсунов. Согласно генеалогии, не только уйсуны, но и род шанышкылы Старшего жуза являются потомками нирун-монголов. Помимо этого, линия C2*-ST идентифицируется у нескольких генеалогических родословных нирун-монголов, как к примеру, у гэнигэсов, мангутов и хатагинов, а также среди хазарейцев. Поэтому в целом исследователи предполагают происхождение Y-хромосомных линий основных популяций Южного Казахстана от нирун-монголов.
Выводы
Результаты исследования представителей Старшего жуза Южного Казахстана показали генетическое сходство шести уйсунских и одного неуйсунского рода друг с другом, в то время как остальные четыре рода (два уйсунских и два неуйсунских) имеют отдельные отцовские линии. Таким образом, генетические данные не во всех деталях воспроизводят традиционную генеалогию казахов. Однако в целом генетические свидетельства согласуются с общим происхождением большинства родов Южного Казахстана.
При этом, значительная часть населения происходит от трёх основателей, которые жили около 700-800 лет назад, в отличие от одного основателя, согласно традиционной генеалогии. Эти три кластера были идентифицированы в пределах гаплогруппы C2*-ST (C-F3796). Первый кластер характерен для большинства уйсунских родов, второй для неуйсунского рода жалайыр Южного Казахстана, а третий-для племени керей в Северном Казахстане, но включает и отдельные образцы из Южного Казахстана. Преобладание гаплогруппы С2*-ST в Южном Казахстане позволяет предположить происхождение большинства Y-хромосомных линий от нирун-монголов.
Исследование также не подтверждает гипотезу происхождения родоплеменного объединения казахов – уйсун, от кочевого племени усуней, которые жили в бассейне реки Тарим, или от дарлекин-монголов.
Источник:
Zhabagin, M., Sabitov, Z., Tarlykov, P. Oleg Balanovsky & Elena Balanovska et al. The medieval Mongolian roots of Y-chromosomal lineages from South Kazakhstan. BMC Genet 21, 87 (2020). doi.org/10.1186/s12863-020-00897-5
Приручение лошади около 5,5 тыс. лет назад представляет собой одно из важнейших технологических достижений древнего мира, поскольку лошади произвели революцию в транспорте и повлияли на модели торговли, войны и миграций. Археологические и органические остатки, а также генетический анализ позволяют предположить, что домашняя лошадь возникла в степях Центральной Азии, после чего распространилась по Восточной Европе, а затем и по Передней Азии. Данные из контекста ботайской культуры Казахстана предполагают, что к середине - концу четвёртого тысячелетия до н. э. лошадей могли содержать и доить. Но эти доводы оспариваются, потому как ряд учёных сочли доказательства одомашнивания лошадей ботайцами не убедительными, хотя и отметили хорошую работу исследователей, но подвергли сомнению интерпретацию результатов. Исследование геномов древних лошадей поставило под сомнение точку зрения о том, что современные домашние лошади произошли из Центральной Азии, поскольку было показано, что так называемые «ботайские» лошади являются предками лошадей Пржевальского в Северо-Восточной Азии, но не являются основным источником древних или современных домашних лошадей. Другое недавнее исследование исключило Пиренейский полуостров, как второй потенциальный центр одомашнивания лошадей, показав, что иберийские дикие лошади вымерли, не оставив заметных следов в геномах современных.
Однако оставалось ещё две области, которые были предложены в качестве центров приручения современных лошадей: это Причерноморско-Каспийская степь и Анатолия. Хотя степь уже долгое время считается наиболее вероятным источником домашних лошадей, Анатолия с этой точки зрения плохо изучена, несмотря на её долгую историю использования диких лошадей, а также репутацию как производителя ценных пород лошадей в античном мире. Комбинация текстов, изображений и археозоологических данных позволяет предположить, что к середине-концу III тысячелетия до н. э. домашние лошади были завезены из соседних горных регионов в Месопотамию (современный Ирак и северо-восток Сирии), где они часто упоминались в клинописных текстах как «горные ослы».

Царь Ассирии Ашшурнацирапал II охотится на львов
Изначально содержавшиеся в небольшом количестве, лошади стали широко известны в Передней Азии в течение нескольких столетий в связи с распространением колесниц, технологического новшества второго тысячелетия до н. э.
Поскольку одомашнивание лошадей в евразийских степях, регионе, исторически известном своими так называемыми “конными культурами”, вероятно, началось в четвёртом или, возможно, даже пятом тысячелетии до н. э., долгое время утверждалось, что лошади Передней Азии являются потомками этих ранних одомашненных степных лошадей, которые прибыли в регион по средствам ещё недостаточно изученных процессов миграций или обмена с участием жителей Причерноморско-Каспийской степи и Кавказских регионов. Всего у лошадей, на данный момент выявлено 18 основных гаплогрупп (от A до R), время разделения которых относится в основном к неолиту и более поздним периодам.
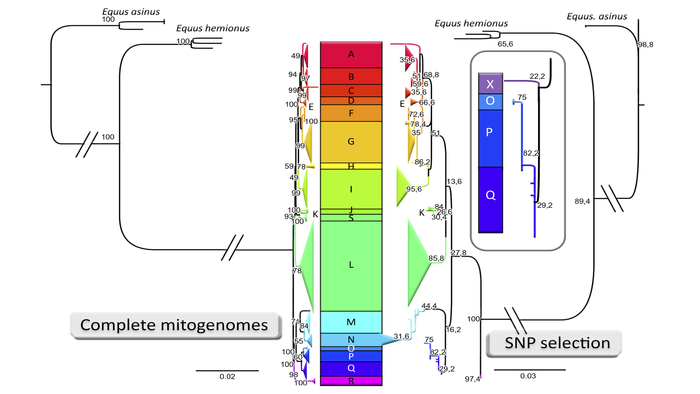
Домашние лошади демонстрируют гораздо более высокую степень генетической изменчивости митохондриальных линий по сравнению с крупным рогатым скотом, а также свиньями и овцами. При этом, большинство митохондриальных линий, наблюдаемых у домашних лошадей, существовало ещё до приручения. Однако эти анализы древних лошадей не дали чёткой филогеографической структуры, которая позволила бы установить пространственно-временное происхождение и центр их одомашнивания. Ведь результаты предполагают, что лошади на севере Евразии постоянно мигрировали и смешивались. А вот что касается мужской линии, то у современных лошадей выявлен только один гаплотип, что привело к поспешным выводам о едином событии одомашнивания. Но анализ древних образцов указал на дополнительные мужские линии в доисторических популяциях до одомашнивания и показал, что генетически разнородные самцы-основатели участвовали в раннем одомашнивании.

Впоследствии это разнообразие сократилось, вероятно, в результате более направленного отбора человеком, который, скорее всего, начался в железном веке и продолжался во времена Римской империи и после VII-IX вв. н. э.
Палеогенетические данные по генетическим локусам, связанным с окрасом шерсти у лошадей, свидетельствуют об увеличении разнообразия окраса шерсти начиная с бронзового века, что считается ранней стадией процесса одомашнивания и является обычным явлением для домашних таксонов по сравнению с их дикими аналогами. Поэтому появление новых окрасов шерсти служит полезным маркером для идентификации домашних лошадей в археологических комплексах.

До сих пор происхождение домашних лошадей в Анатолии оставалось малоизученным, однако тщательное извлечение останков лошадей из хорошо датированных археологических контекстов в Анатолии и на соседнем Кавказе, совместно с прогрессом в палеогенетических методах, теперь позволяет конкретно рассмотреть процессы, ответственные за происхождение домашних лошадей в этой части Передней Азии.
В своей работе международная группа исследователей объединила морфологическую классификацию останков непарнокопытных с палеогенетическим анализом митохондриальной ДНК, Y-хромосомы и аутосомных ДНК-маркеров, связанных с окрасом шерсти, для отслеживания пространственно-временной динамики появления домашних лошадей в Анатолии.
Чтобы прояснить давний вопрос в истории Ближнего Востока о местном приручении лошадей в Анатолии было проанализировано более 100 останков лошадей из 14 доисторических поселений Центральной Анатолии и Кавказа, охватывающих большую часть голоцена, а именно с 9-го тысячелетия до н. э. по 1-е тысячелетие н. э.
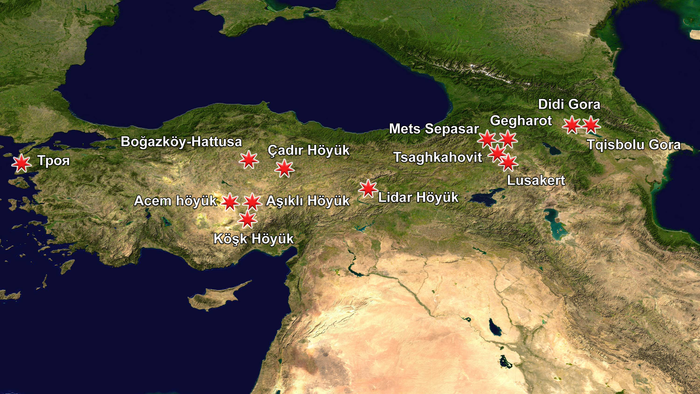
Результаты
Дикие и домашние лошади в Анатолии
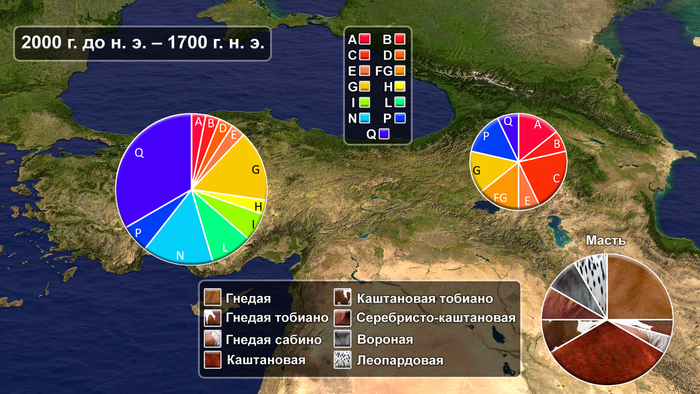
Результаты позволяют сделать вывод о том, что домашние лошади были завезены на Кавказ и в Анатолию по крайней мере в 2000 г. до н. э., предположительно из Евразийской степи.
Этот вывод основан на том факте, что в Анатолии местные популяции лошадей до 6,5 тыс. лет назад несут только две митохондриальные гаплогруппы, P и X, причём последняя является ранее незарегистрированным гаплотипом, принадлежащим поддереву O-P-Q. До сих пор эти гаплогруппы не встречались где-либо ещё в Евразии в современном или более раннем контексте. Более того, гаплотип X, вероятно, имел ограниченное временное распространение в Анатолии голоцена, возможно, исчезнув после 5500 г. до н.э.
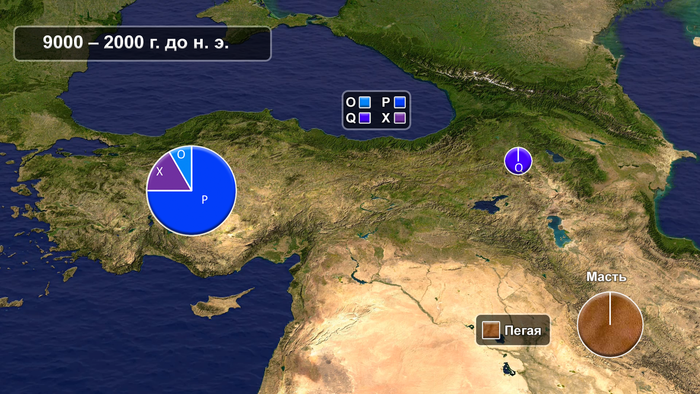
Данные подтверждают вывод о том, что эти две гаплогруппы отражают локальные митохондриальные признаки диких лошадей, на которых охотились в Анатолии в раннем и среднем голоцене. Исследователи предполагают, что гаплогруппы P и X развивались независимо в Анатолии в течение позднего плейстоцена и раннего голоцена со слабым или полностью отсутствующим потоком генов от соседних популяций диких лошадей из-за географических барьеров, отделяющих Анатолию от северной Евразии, а именно Босфора, а также гор Загрос и Кавказа. Таким образом, новое исследование предоставляет первые доказательства того, что Анатолия была домом для генетически отличающейся популяции диких лошадей, которые, согласно археозоологическим находкам, широко использовались в периоды неолита и энеолита. Остатки лошадей раннего неолита Ашиклы-Хююк, неолита / энеолита Кёшка-Хююк и позднего энеолита / раннего бронзового века Чадир-Хююк, являются представителями этих диких анатолийских лошадей.
А примерно в 2000 г. до н.э. наблюдается статистически значимое снижение частот местных митохондриальных линий диких лошадей, поскольку гаплогруппа P становится редкой, а гаплотип X полностью исчезает. Хотя низкая частота гаплотипа X у лошадей отмечена ещё до бронзового века.
Параллельно с этим, разнообразие материнских линий у лошадей Кавказа и Анатолии заметно возросло от 2 до 14, т.е. уровня современных, а также лошадей эпохи энеолита, халколита и ранней бронзы из Юго-Восточной Европы и Казахстана.
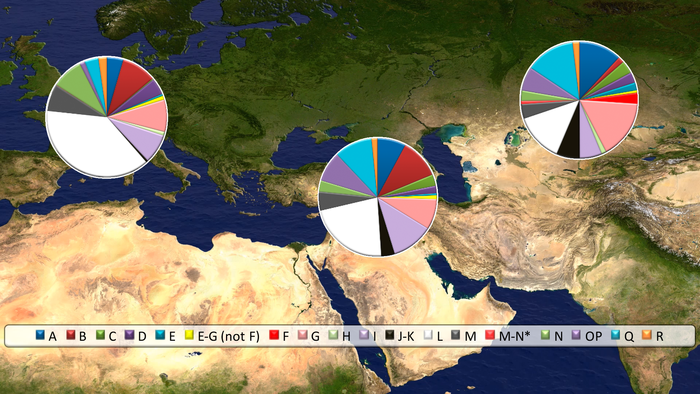
В исследованиях лошадей Евразийской степи, митохондриальные гаплогруппы не демонстрируют филогеографической структуры в Евразии, что согласуется с отсутствием значительных физических барьеров в обширных евразийских степях. Это предполагает, что у популяций лошадей степи был постоянный обмен генетической информацией между её членами, что, вероятно, объясняет высокое разнообразие современных популяций домашних лошадей. Это также объясняет быстрый рост разнообразия, наблюдаемый в наборе данных из этого исследования при внедрении лошадей на Кавказ и в Анатолию. Это внезапное появление неместных родословных совпадает с появлением иконографических и эпиграфических свидетельств о лошадях и верховой езде в конце третьего тысячелетия до нашей эры и свидетельствует о значительном импорте домашних форм и, следовательно, против независимого местного процесса одомашнивания. Результаты, полученные при генотипировании аллелей, связанных с вариациями в цвете шерсти, подтверждают этот вывод, поскольку лошади бронзового века в Анатолии и на Кавказе демонстрируют мутации, соответствующие вариантам окраски шерсти, которые, как считается, были выбраны во время одомашнивания в евразийских степях, например, каштановый, чёрный и серебристый.
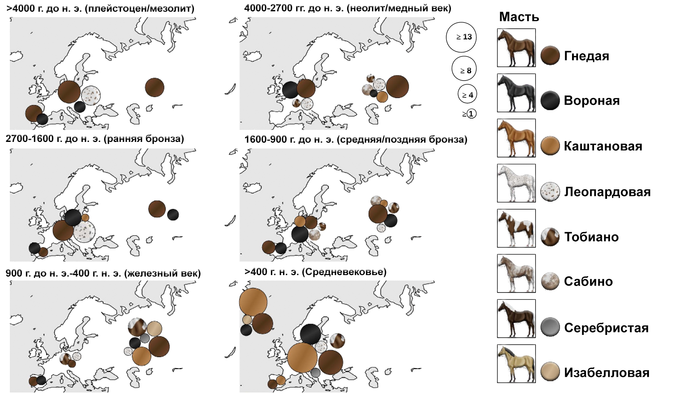
А мутации, связанные с разбавлением цвета шерсти или пятнами, появились позже, после 1200 г. до н.э. согласно данным исследования. Таким образом, исходя из новых генетических данных, исследователи делают вывод, что домашние лошади, завезённые в Анатолию в бронзовом веке, несли мутации, обнаруженные ранее в Северной Евразии и, следовательно, полученные преимущественно от лошадей из этого обширного региона. Хотя окончательное географическое происхождение этой неместной популяции невозможно определить с помощью имеющихся данных, евразийские степи к северу от Чёрного моря кажутся наиболее вероятным кандидатом. Однако местный анатолийский гаплотип P сохранялся у домашних лошадей бронзового века Анатолии и Кавказа, а также у современных лошадей, правда с низкой частотой, около 8%. А это позволяет предположить, что дикие анатолийские лошади, вероятно, часто включались в стада домашних, вскоре после появления их регионе и исчезновения в дикой природе. Результаты свидетельствуют о быстром переходе от охоты к скотоводству после появления домашних лошадей в раннем бронзовом веке, что, вероятно, коррелирует с окончательным сокращением популяции диких лошадей в Анатолии и на Кавказе, на что указывает низкая частота их останков. Схожая картина с двумя гаплотипами Y-хромосомы, Y-HT-1 и Y-HT-3, Анатолии и Кавказа, намекает на возможность популяционной динамики, подобной той, что зафиксирована для Северной Евразии, где преобладающий Y-гаплотип –Y-HT-1 у современных лошадей, заметно увеличился после начала одомашнивания, достигнув фиксации в генофонде к средневековью, в то время как Y-HT-3 снижался с течением времени вплоть до своего исчезновения.
В Анатолийском наборе данных из этой работы оба гаплотипа встречались с сопоставимой частотой около 2000 года до н. э., но и здесь частота Y-HT-3 со временем уменьшилась, а самая последняя лошадь, которая несла Y-HT-3, была родом с Кавказа, возрастом около 3300 лет. Это уменьшение разнообразия Y-хромосом, по-видимому, является результатом сильного отбора жеребцов.
Поскольку северная Евразия, и в частности Причерноморско-Каспийская степь, в настоящее время является наиболее вероятным местом происхождения домашних лошадей, завезённых в Анатолию, существует два возможных маршрута: один через Юго-Восточную Европу и один через Кавказ. Маршрут через Босфор был предложен на основе самых ранних зооархеологических свидетельств существования домашних лошадей на южных Балканах на стоянке раннего бронзового века Канлигечит около 2600–2300 гг. до н. э. Окрасы шерсти 10 лошадей с этого участка были генотипированы и выявили сильно смещённое распределение мутаций окраса шерсти у 6 из 10 гомозиготных черных лошадей, 4 из которых также показывают "пятнистость леопарда" ещё у 2 гнедых лошадей с пятнистостью, мутаций каштанового цвета не обнаружено. Эта картина сильно контрастирует с новыми данными из Анатолии и Кавказа, где каштановый цвет является самым ранним вариантом окраски шерсти, в то время как черные и леопардовые окрасы остаются очень редкими (только 2 из 25).
Эти различия служат аргументом против введения популяции домашних лошадей, аналогичной той, что была обнаружена в Канлигечите. Более того, нет никаких археологических данных о коневодстве в Западной Анатолии в третьем тысячелетии до нашей эры, что не даёт дополнительной поддержки гипотезе о раннем вводном пути через Босфор.
Напротив, новая идентификация нескольких неместных митохондриальных линий и мутаций, связанных с окрасом шерсти, широко появляющихся одновременно на Южном Кавказе и в центральной Анатолии, свидетельствует в пользу пути распространения через Кавказ. Наличие костей и изображений лошадей в поселениях майкопской культуры и погребениях около 3300 г. до н.э. на Северном Кавказе привело к предположению, что практика верховой езды началась в майкопский период. Кроме того, недавние исследования геномов людей того же периода показали непрерывный поток генов между степями медного века и народами Кавказа, а позже, в эпоху бронзы, между Месопотамией, Анатолией, Южным и Северным Кавказом, а также степью. Этот обмен между группами людей, похоже, усиливается с началом похолодания и засухи 4200 лет назад, что могло повлиять на стратегии выживания и социальные сети в степной зоне.
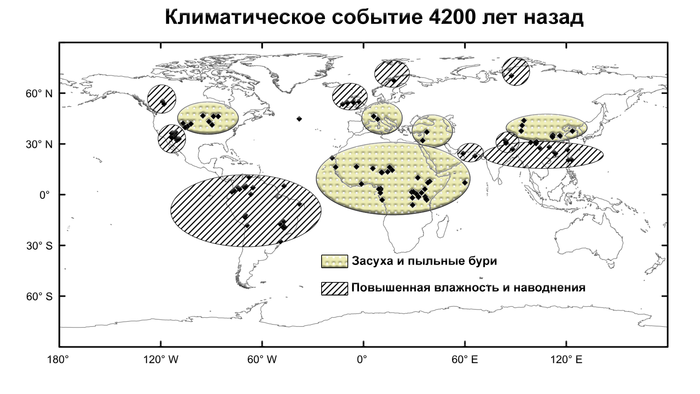
Согласно имеющимся данным, это климатическое событие, по-видимому, в значительной степени совпало с появлением неместных митохондриальных гаплогрупп и сменой окраса шерсти лошадей на Кавказе и в Анатолии. Что также связано с распространением коневодства и, возможно, индоевропейских языков. Хотя культурные процессы, инициировавшие распространение коневодства к югу от Кавказа, в настоящее время трудно поддаются анализу, они могут быть связаны с перемещением людей через Кавказ в Анатолию, начиная с конца третьего тысячелетия до нашей эры.
Древний мул
Среди образцов из новой работы попался и мул, а именно потомок лошади и осла, из контекста раннего железного века, в Чадир-Хююк Центральной Анатолии, датируемого 1100–800 гг. до н. э. Из данного региона это первое свидетельство мулов, помимо европейского железного века и римского периода. При этом, учитывая, что гаплогруппа кобылы была неместной, получается, что гибрид был от домашних животных. При этом кобыла передала мулу два мутантных аллеля цвета шерсти. Хотя это и не является чем-то удивительным, присутствие мула в железном веке Анатолии отражает новую роль домашних лошадей, появившуюся по мере их распространения в регионе, где домашние ослы использовались с четвёртого тысячелетия до нашей эры. Однако этот случай является ранним примером преднамеренной гибридизации. Будучи самым дорогим видом, упомянутым в ценах на домашний скот хеттского периода от ~ 1600 до ~ 1178 г. до н. э., мулы, вероятно, ценились очень высоко.
Помимо лошадей, результаты нового исследования недвусмысленно свидетельствуют о том, что люди неолита, энеолита и бронзового века Анатолии также охотились на европейских плейстоценовых ослов, которые когда-то населяли большую часть Анатолии. Таким образом, объединённые данные предполагают, что ослы вымерли в Анатолии в эпоху поздней бронзы более или менее одновременно с местными дикими лошадьми, возможно, в ответ на ту же комбинацию факторов, включая повышенную засушливость, связанную с засухой 2100-х годов до н. э., а также конкуренцию за пастбищные ресурсы с растущим количеством скота и, вероятно, из-за охоты элит.
Выводы
В новой работе, охватывающей ~ 9000 лет голоцена, были проанализированы митохондриальные линии древних лошадей из Южного Кавказа и Анатолии. Исследование позволило проверить гипотезу о том, что Анатолия была центром одомашнивания лошадей. Авторам работы удалось идентифицировать митотипы, характерные для местных диких лошадей Анатолии, которые регулярно использовались в раннем и среднем голоцене. При этом генетический анализ показал, что неместные генетические маркеры, которые все ещё присутствуют у домашних лошадей, появились в регионе не постепенно, а внезапно после ~ 2000 г. до н.э. Исследование также демонстрирует, что у этих завезённых домашних лошадей окрас отличался от местных до приручения. Однако материнская линия P, выявленная у местных лошадей до приручения, присутствует и в бронзовом веке, что подразумевает ограниченное включение местных кобыл в домашние стада.
В целом результаты указывают на то, что домашние лошади были завезены в Анатолию, возможно, через Кавказ в период бронзового века, и дают дату начала эксплуатации домашних лошадей в Анатолии и Закавказье около 2000 года до н. э. Данные также свидетельствуют против местного независимого приручения лошади в этом регионе. Новые результаты убедительно указывают на то, что Анатолия не была основным источником происхождения домашних лошадей, но, как наблюдалось и в других регионах, местные материнские линии всё же были включены в стада неместных домашних лошадей, которые также были скрещены с местными ослами для создания мулов. Однако точное географическое происхождение завезённых лошадей ещё предстоит определить.
Источник: Ancient DNA shows domestic horses were introduced in the southern Caucasus and Anatolia during the Bronze Age
Silvia Guimaraes, Benjamin S. Arbuckle, Joris Peters, Sarah E. Adcock, Hijlke Buitenhuis, Hannah Chazin, Ninna Manaseryan, Hans-Peter Uerpmann, Thierry Grange, and Eva-Maria Geigl doi.org/10.1126/sciadv.abb0030
Хунну представляют собой объединение кочевых скотоводческих племён и не только, которые основали первую степную империю во Внутренней Азии. С III века до н. э. и по II век н. э. они правили территорией, простиравшейся на своём пике от озера Байкал на севере до пустыни Гоби на юге и от Западной Маньчжурии на востоке до Памира на Западе.

А их сложные отношения с соседней империей Хань, привели к строительству Великой Китайской стены в ответ на набеги хунну. Но также были периоды торговых отношений, даров, дани и браков по договорённости, что входило в политику “гармоничных отношений”, принятой в империи Хань. Однако около 130 года до н. э. китайцы начали проводить более агрессивную политику, что привело к упадку династии хунну, упоминания о которой исчезли из китайских записей после II века н. э.
Поскольку хунну не оставили письменных источников, большая часть доступной информации об их образе жизни, социальных, экономических и политических системах, а также о военных практиках, содержится в китайских исторических текстах. При этом ввиду географического положения границы с Китаем, больше информации присутствует о хунну из южных регионов. И хотя археологические раскопки были ещё одним важным источником данных, некоторые ключевые аспекты культуры хунну до сих пор не ясны, а именно их биологическая структура, этническая или языковая идентичность и их связь с другими кочевниками железного века. Термин "хунну" обозначал как народ, так и политическое образование, но неясно, может ли он быть отнесён к этнической группе, биологической популяции, археологической культуре или, что более вероятно, к какой-то их комбинации.
Результаты физико-антропологических исследований и особенно сравнительного черепно-лицевого анализа выявили сложную популяционную структуру населения империи Хунну, преимущественно как смесь между европеоидными и монголоидными этническими группами, несмотря на преобладающую монголоидную внешность.

А генетический анализ скелетных останков, обнаруженных в погребениях эпохи хунну, также выявил черты, характерные для восточноазиатских популяций, с примесью западноевразийских.

Проще говоря, хунну это объединение различных племён, населявших обширные территории будущей империи, причём эти люди говорили на разных языках и представляли предков различных современных этнических групп. Например, представители хунну из Западной и Центральной Монголии по данным физической антропологии близки к культурам тюркского круга, а Восточной Монголии к сяньби и хунну Забайкалья. Существует ряд теорий и мнений, по поводу языковой принадлежности хунну, но интересно, что самые ранние хуннские имена II века до нашей эры имели иранский тип, отсюда и предположения о близости к скифам сакам, однако часть хунну можно рассматривать также как прототюрков и носителей енисейских языков. С большой долей вероятности это был конгломерат как культурный, так и языковой.
В новой работе были получены данные ДНК из скелетных останков 52 человек, найденных на кладбище Тамир Улан Хошуу в Центральной Монголии. Это кладбище использовалось в середине периода хунну с I века до н. э. по I века н. э. Тамир Улан Хошуу расположено недалеко от слияния реки Тамирын-Гол и реки Орхон в аймаке Архангай Центральной Монголии. Кладбище занимает территорию в 22 гектара и включает в себя в общей сложности 397 могил, ограниченных каменными кругами. Участок разделён оврагом на западную часть, состоящую из 344 могил, и восточную часть, состоящую из 53 больших могил (до 20 м в диаметре), отражающих стиль гробниц, типичный для эпохи хунну и, возможно, принадлежащих элите.

Хотя почти все гробницы были разграблены, в захоронениях было найдено много погребального инвентаря, в том числе золотые серьги, стеклянные бусы, керамические кувшины, железные ножи, лаковое дерево, а также импортированные из Китая артефакты, такие как фрагменты бронзовых зеркал и круглые нефритовые камни. Помимо этого, орнаменты в виде четырёхлистника, характерные для культуры хунну, были обнаружены на нескольких гробах. Как и в других гробницах хунну, рядом с умершими, вне гробов, были захоронены головы и конечностей овец, лошадей, а также быков.
В 48 раскопанных могилах были обнаружены останки 57 человек, из них 21 мужчина, 17 женщин и у 19 человек пол определить не удалось, однако далее в этом деле помогла генетика. Хотя большинство могил были одиночными, в семи были останки двух или трёх человек. А четыре могилы были отнесены к культурам тюркского круга.

Чтобы получить более полное представление о народах, которые составляли группы, известные как хунну, и получить представление об их происхождении и потенциальных биологических отношениях с другими кочевыми группами, авторы работы исследовали генетическую структуру и родство людей, похороненных в Тамир Улан Хошуу.
С помощью комбинированного анализа ДНК-маркеров, расположенных на аутосомах, Y-хромосоме и целых митохондриальных геномах, генетики реконструировали беспрецедентную древнюю семейную структуру хунну, вероятно, элитную семью из 19 человек и пяти поколений.
Также авторы подчёркивают, что в работе были исследованы маркеры мтДНК и Y-хромосомы не на уровне гаплогрупп, а на уровне гаплотипов или маркеров коротких тандемных повторов, что даёт более точный результат в попытке распутать генетическую историю хунну.
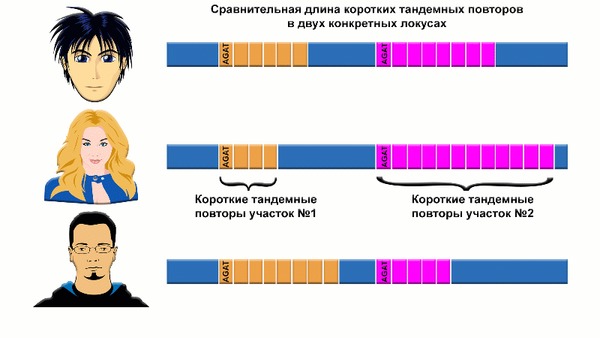
Анализ коротких тандемных повторов не разрезает ДНК рестрикционными ферментами или своеобразными биохимическими ножницами, вместо этого полимеразная цепная реакция используется для определения длины коротких тандемных повторов, путём образования дополнительных копий участков.

Эта длинна у каждого человека уникальна и зависит от количества повторов на каждом участке. Метод широко используется в судебно-медицинской экспертизе и стал доступен в связи с хорошей сохранностью останков 52-х из 57-и человек.
Результаты
Родственные связи
Благодаря молекулярной идентификации пола удалось выявить 26 мужчин и 26 женщин, что соответствовало доступным физико-антропологическим данным. Попарное сравнение и тест отношения правдоподобия выявили наличие родственников первой и второй степени. Было выделено 5 нуклеарных семей, состоящих из родителей и одного или двух детей. Все они были частью семейной структуры из пяти поколений, состоящей из 19 человек.

За пределами этой семьи было также идентифицировано 5 пар родитель / ребёнок, которые были похоронены вместе или близко друг к другу.
Y-гаплотипы, состоящие из 19-27 коротких тандемных повторов были получены для всех 26 исследованных мужчин, из них можно было выделить шестнадцать различных Y-гаплотипов, 6 из которых были общими для 2-5 человек.
А 28 различных гаплотипов мтДНК, 10 из которых были общими для 2-8 человек, также подтвердили и уточнили родственные отношения захороненных.
Гаплогруппы Y-хромосомы и сравнение гаплотипов
У проанализированных людей определены субклады 5 основных гаплогрупп Y-хромосомы, а именно R, Q, N, J и G.
10 человек, имели субклады R1a Z2125 и 4 человека Z95. Пять человек были отнесены к субкладу Q1a-M120. У двух человек была определена гаплогруппа N, а ещё у двух гаплогруппы J1 и G1. Последняя определена с вероятностью всего 77,6%, также не определены судклады гаплогрупп N, J1 и G1.

А вот теперь перейдём к самому интересному...
Y-гаплотипы были индивидуально сопоставлены с внутренней базой данных коротких тандемных повторов. Сразу отмечу, что чем больше совпадений, тем ближе исследованные люди к друг. Этот самый высокий процент совпадений был получен для представителей хунну, с гаплогруппой R1a. Большинство из них имели Y-гаплотипы, близкие к таковым у мужчин железного века из Красноярского края, намекая на сибирских скифов.
Наиболее близкие гаплотипы наблюдались между образцами Тамир 09A и 10 и двумя представителями тагарской культуры республики Хакасия.
Совпадения наблюдались также с мужчинами среднего бронзового века той же местности, к примеру вышеупомянутый образец Тамир 10, разделял схожие гаплотипы с двумя представителями андроновской культуры. Близкие совпадения были обнаружены также с 2 представителями скифо-сибирского мира из Республики Тыва. Помимо этого, Тамир 09A и 10 продемонстрировали общий гаплотип с венгерским королём из династии Арпадов — Белом III. При этом образцы хунну Тамир 12 и 35 показали схожесть с другим мужчиной, из королевской базилики Секешфехервара, где был похоронен король Бела III (1172-1196 гг.). Интересно, что недавнее исследование установило, что ближайшими живыми родственниками династии Арпадов являются современные башкиры преимущественно из Бурзянского и Абзелиловского районов Республики Башкортостан Российской Федерации.

А происхождение первой венгерской королевской династии прослеживается до регионов Северного Афганистана около 4500 лет назад и идентифицирует башкир как их ближайших родственников, с датой разделения между двумя популяциями в начале первого тысячелетия нашей эры. Т.е. популяции ведущие к современным башкирам и к Арпадам разделились около 2000 лет назад. Это предполагает, что группа людей, состоящая из носителей гаплогрупп ассоциированных с тюрками R1a-SUR51 и финно-угорами N-B539, покинула Волго-Уральский регион около 2000 лет назад и начала миграцию, которая в конечном итоге завершилась поселением в Карпатском бассейне.
Но вернёмся к хунну...
Один из вышеупомянутых представителей, Тамир 12, разделяет по меньшей мере восемь коротких тандемных повторов с хунну из поселения в долине реки Эгийн-Гол Монголии. Двое мужчин со множеством совпадений Тамир 9A и 10, совместно с ещё двумя хунну Тамир 18 и 24 несли гаплотипы, как у венгерских завоевателей (мадьяр) X века. А представители Тамир 36 и 48 могли быть связаны с простым воином, найденным в могилах Таван-Толгой (1130-1250 гг. н. э.) монгольской императорской семьи.
Примечательно, что Тамир 13А, отнесённый к представителям тюркского круга по материальной культуре и нёсший гаплогруппу Y-хромосомы N, показал некоторое сходство с представителями Центральной Якутии XVIII века, а также чуть меньшее с аварами VII–VIII веков и венгерским завоевателем X века.
Для других проанализированных людей с кладбища Тамир Улан Хошуу не выявлено совпадений, но это не значит, что из нет, просто база древних гаплотипов слишком мала (~ 216 000 гаплотипов).
Гаплогруппы мтДНК и сравнение гаплотипов
Идентифицированные 28 различных гаплотипов были отнесены к 11 различным основным гаплогруппам ассоциированных как с западноевразийским происхождением (H, J, T, U, X), так и с восточноевразийским (A, B, C, D, F, G).

Для оценки географического и временного распределения этих материнских линий было проведено сравнение целых последовательностей митохондриальной ДНК с последовательностями древних и современных людей.
Западноевразийские линии
Наиболее распространённый гаплотип (№1), обнаруженный у 7 из 19 членов большой семьи и у одного неродственного представителя Тамир 35, был связан с гаплогруппой мтДНК T2b3+151 и показал наиболее близкие совпадения (от 3 до 4 различий) с людьми, связанными с комплексом колоколовидных кубков во Франции и Испании. Другие материнские линии, выявленные в семье, относятся к гаплогруппам J2a1a1 (№2), J2b1a2a (№15) или X2b4a (№10). Эти линии также были близки к тем, которые встречаются у представителей колоколовидных кубков или срубной культурно-историческая общности Восточной Европы. Таким образом, все материнские линии, обнаруженные в выявленной большой семье, имели западноевразийское происхождение.
Два гаплотипа (№3 и №16), принадлежащих к гаплогруппе T1a1, были близки к людям, ассоциированным с комплексом колоколовидных кубков из Чехии, Германии и Великобритании, а также к средневековому жителю из Венгрии, связанного с лангобардами. Второй наиболее представленный гаплотип (№13), обнаруженный у пяти членов семейной группы из 5 поколений и связанный с гаплогруппой H47, довольно отдалённо связан (5 различий) с человеком из неолитической стоянки в Восточной Венгрии. Ещё два гаплотипа хунну, принадлежащих гаплогруппам H5a1 и H7b (№7 и №23) были близки к представителям неолита и бронзового века Европы (4 различия).
Другие европейские линии, принадлежали гаплогруппам H2a1 (№21), J1d5 (№12), U2e2a1 (№14), U5a1a1 (№24) и U5a1+a1 (№27). Но из них только последние два гаплотипа были тесно связаны (от 2 до 3 различий) с обнаруженными у представителей культуры колоколовидных кубков. Помимо этого, у мужчины Тамир 44, с гаплогруппой мтДНК U5a1+a1, были совпадения (4 различия) с представителями Баденской культуры, лангобардов VI-VII веков и аварских групп.
Восточноевразийские линии
Что касается восточноевразийских линий, то совпадений оказалось не так много, как в случае с западноевразийскими, вероятно в связи с меньшим числом опубликованных данных от образцов из азиатских археологических памятников.
Тем не менее, гаплотип мтДНК №22, переносимый Тамир 37 и связанный с гаплогруппой D4j8, имел 4 различия с гаплотипом представителя Якутии XIX века. А гаплотип Тамир 46, связанный с гаплогруппой D4j7 был тесно связан с гаплотипами 3 человек скифо-сибирского мира из Республики Тыва (также 4 различия). Гаплотип (№19) Тамир 30B показал всего 3 различия с гаплотипом захороненного в позднесарматских Темясовских курганах Республики Башкортостан I-III века. И гаплотип №26, гаплогруппы G2a1d2 образцов Тамир 43 и 45 продемонстрировал единственное различие с венгерским завоевателем X века.
Фенотипический анализ членов семьи
Фенотипический анализ 7 из 19 представителей большой семьи позволил оценить их цвет волос, глаз, а также биогеографическую родословную.

Образец Тамир 01, который разделял европейскую линию мтДНК T2b3+151 со своей родной или сводной сестрой Тамир 15 и её потомством, нёс азиатский гаплотип гаплогруппы Y-хромосомы Q1a-M120 и имел черные волосы и карие глаза, а также смешанное европейско-восточноазиатское происхождение, впрочем, как и ожидалось. В свою очередь Тамир 01 вступил в контакт с женщиной Тамир 02C которая по материнской линии была европейского происхождения с гаплогруппой J2a1a1. Их дети, Тамир 03 и Тамир 26, имели уже более смешанное происхождение и цвет их волос, по-видимому, был немного светлее. Далее Тамир 26 завёл потомство с двумя женщинами, также с западноевразийскими материнскими линии, жёнами или нет не известно, были найдены только потомки, при чём все женского пола, несущие гаплогруппу H47. Одна из дочерей, Тамир 19, вышла замуж за Тамир 23 с отцовской линией R1a Z95 и материнской линией J2b1a2a тоже европейского происхождения. У их сына Тамир 18 были более светлые волосы, а по результатам, генетически он уже был полностью европейского происхождения, но родившийся на территории современной Монголии. Также прогноз европейского генетического происхождения для Тамир 25, внука Тамир 15, согласуется с его отцовскими и материнскими западноевразийскими гаплотипами, принадлежащими гаплогруппам R1a-Z2125 и T2b3 соответственно.
Итоги
Совместный анализ аутосомных и однополых маркеров показал, что участок Тамир Улан Хошуу представлял собой семейное кладбище без признаков различий по полу. Мужчины и женщины были захоронены в равных соотношениях, в одних и тех же типах гробниц и со схожим количеством погребального инвентаря. Подобная практика наблюдалась и на другом кладбище хунну в Северном Китае – могильник Даодуньцзы.
Было высказано предположение, что такая практика, вероятно, была присуща кочевым скотоводам, чьи средства к существованию зависят как от мужчин, так и от женщин и детей.
Это контрастирует с другими кладбищами, где отсутствуют взрослые потомки женского пола, что объясняется женской экзогамией, т.е. заключением ими браков за пределами своей социальной группы.
Многомаркерный подход позволил реконструировать генеалогическое древо, охватывающее пять поколений с двумя ветвями. Первая ветвь состояла из потомков мужчины Тамир 01, умершего в возрасте более 30 лет, а вторая ветвь из потомков его родной или сводной сестры Тамир 15, которая была похоронена в возрасте более 50 лет, в одной из самых больших и глубоких могил южной части кладбища. Они унаследовали западноевразийскую материнскую линию T2b3, а брат Тамир 01 в добавок унаследовал восточноевразийскую отцовскую линию Q1a M120. Носители этой гаплогруппы Y-хромосомы, по-видимому, мигрировали из Монголии в Китай в период неолита и распространились по Китаю около 3 тыс. лет назад. Гаплогруппа Q1a M120 наблюдалась с высокой частотой у жителей Центральной равнины Китая — Чжун Юань. Исследователи предполагают, что обнаруженная семья из пяти поколений берет своё начало от смешанных браков хунну европейско-восточноазиатского происхождения, вероятно, это было пристижно. При этом потомки этого союза образовывали последующие пары с мужчинами или женщинами с линиями западноевразийского происхождения. Помимо образцов Тамир 01 и его сына Тамир 26, все члены этой предполагаемой элитной семьи имели западноевразийские материнские линии H, J, T, X и отцовскую линию R1a. В связи с чем, потомки Тамир 01 в каждом последующем поколении, а именно, дети Тамир 03 и 26, а также его внук Тамир 23 и правнук Тамир 18 имели всё более светлые волосы.
Проще говоря, у хунну смешанного европейско-восточноазиатского происхождения, после заключения браков с людьми, у которых было больше западноевразийской родословной, в каждом последующем поколении рождались дети с генетическим профилем всё ближе и ближе к европейскому. К примеру, у представителя хунну смешанного происхождения Тамир 01, внук Тамир 23 и правнук Тамир 18 показывают уже полностью европейский генетический профиль. Обратите внимание, какие изменения происходят всего за 3 поколения. Примечательно, что потомки женщины Тамир 15 и их супруги (Тамир 36, 34, 48, 25, 13Б и 40) были найдены в самых крупных захоронениях, что является маркером высокого социального статуса. Это может свидетельствовать о том, что после смерти её родного или сводного брата Тамир 01, Тамир 15 могла стать лидером этой семьи. А что стало тому причиной, либо его некомпетентность, либо даже азиатское происхождение по отцовской линии, как упоминают авторы, установить пока не получается.
Описание гробницы статусной женщины на кладбище Сигупань периода хунну, поддерживает гипотезу о высоком статусе женщин и предполагает, как и некоторые предыдущие исследования, что у кочевников раннего железного века, женщина могла также достичь высокого положения в обществе, как и мужчина. Возможно, что, передавая власть своим дочерям, вожди того периода были уверены в сохранении полного контроля над своими территориями.
Помимо этого, Тамир 24, также похороненный в одной из крупнейших гробниц некрополя, был обозначен как возможный родственник дочери Тамир 15 – Тамир 34, а это усиливает мнение о том, что эта ветвь генеалогии связана с особенно высоким статусом.
А в целом анализ мест захоронения показал, что решения о похоронах основаны на сочетании семейных связей и статуса. Мужей и жён хоронили близко друг к другу, а также к их детям, умершим в подростковом возрасте или младенчестве, в отличие от взрослых детей с собственными семьями, у которых были отдалённые могилы. Если ребёнок умирал раньше своих родителей, он мог быть похоронен рядом с предком, как к примеру, Тамир 11 была похоронена рядом со своей прапрабабушкой Тамир 15. Подобные практики захоронения, на основе биологического родства, также были описаны, хотя и в меньшей степени, в некрополе хунну в долине реки Эгийн-Гол Северной Монголии.
Генетические сравнения образцов Тамир Улан Хошуу с другими группами хунну не выявили большого сходства в составе Y-гаплогруппы, невзирая на то, что R1a1a ранее была зарегистрирована в захоронении элиты хунну в Дуурлинг Нарз в Северо-Восточной Монголии и у трёх человек долины Эгийн-Гол.
Среди новых образцов все гаплотипы R1a тесно связаны и были отнесены к возникшей в евразийских степях азиатской ветви гаплогруппы R1a-Z93, а именно к двум субкладам Z95 или Z2125.

Большинство из этих гаплотипов были очень близки к таковым у южносибирских жителей среднего бронзового или железного века из Красноярского края и Республики Тыва. Что указывает на наличие среди предков хунну представителей андроновской культуры и сибирских скифов, как это и предполагалось ранее, намекая на енисейскую гипотезу. Кроме того, эти гаплотипы совпали с таковыми у мадьяр, что может указывать на сохранение некоторых отцовских линий хунну в генофонде ранних венгерских завоевателей.
Помимо этого, совпадения на уровне гаплотипов наблюдались между ассоциированным с тюрками носителем гаплогруппа Y-хромосомы N – Тамир 13A и аварами VII века, а также с якутами XV-XIX веков. Этот, так называемый, тюркский гаплотип, связанный с гаплогруппой N1a1 или N-Tat, может указывать на то, что авары произошли от людей, живших в Монголии и в районе озера Байкал. Это могло также стать первым прямым доказательством того, что тюркоязычные якуты были сформированы при участии людей из той же местности.
Исследованные представители элиты хунну демонстрируют более высокое разнообразие материнских линий, в противовес отцовским, что согласуется с патрилокальными браками, когда жена переходит в общину или в дом мужа, поскольку единственными взрослыми женщинами в генеалогии были жены, похороненные рядом со своими мужьями. Однако заметными исключениями были статусная женщина Тамир 15, которая была похоронена на том же кладбище, что и её брат, а также её дочь Тамир 34, которая была похоронена на том же кладбище, что и её мать, несмотря на то, что у неё самой была семья и дети.
Разнообразный митохондриальный генофонд Тамир Улан Хошуу содержит равные пропорции западных и восточных компонентов, контрастируя с некрополем Эгийн-гол, в котором большинство гаплогрупп мтДНК имели азиатское происхождение.
Этот баланс между востоком и западом в линиях мтДНК Тамир Улан Хошуу в основном обусловлен генетическим вкладом большой семьи, в которой все материнские линии имели западноевразийское происхождение, в то время как две трети людей за пределами этой семьи имели восточноевразийское генетическое происхождение. Наиболее похожие гаплотипы мтДНК встречаются к западу от Урала, главным образом у людей, связанных с комплексом колоколовидных кубков, что не удивительно, учитывая их контакты со степными популяциями. А некоторые совпадения по мужской линии наблюдались также с раннесредневековыми аварами и мадьярами
В работе прошлого года было отмечено, что восточноевразийские субклады R1a-Z94 и R1a-Z2124 были общими для элиты гуннов, аваров и мадьяр, и, скорее всего, принадлежали к ветви, которая наблюдалась у представителей хунну этого исследования.

Более того, гаплогруппы Q1a и N1a также были основными компонентами этих кочевых групп, что усиливает мнение о том, что гунны, а следовательно, авары и венгерские завоеватели — мадьяры, могли в значительной степени происходить от хунну.
После того как хунну были побеждены и поглощены китайской династией Хань, их северные представители бежали на северо-запад, а уже часть их потомков, вероятно, мигрировала через Евразию и завоевала Карпатский бассейн. А другая часть хунну участвовала в формировании якутских популяций, на что указывают гаплотипы Тамир Улан Хошуу.
Подводя окончательный итог следует отметить, что это обширное исследование захоронений хунну позволило исследователям реконструировать большую семью из 5 поколений и показать, что племена хунну Центральной Монголии хоронили своих умерших в соответствии с семейными связями. При этом результаты показали, что по крайней мере некоторые женщины могли быть социально равными мужчинам. И, пожалуй, для многих зрителей будет самым интересным то, что у хунну прослеживаются предковые линии сибирских скифов, а некоторые отцовские и материнские гаплотипы самих хунну обнаружены в генофонде гуннов, аваров, а также мадьяр. Т.е. народов скифо-сибирского происхождения, которые ассоциируются с завоеваниями...
Источник: Keyser, C., Zvénigorosky, V., Gonzalez, A. et al. Genetic evidence suggests a sense of family, parity and conquest in the Xiongnu Iron Age nomads of Mongolia. Hum Genet (2020). doi.org/10.1007/s00439-020-02209-4
В начале эпохи викингов, с VIII века, вдоль берегов Северного и Балтийского морей появились новые поселения, которые функционировали как центры морской торговли или эмпории, например, Рибе, Каупанг, Бирка, Хедебю, Рерик и Охус.
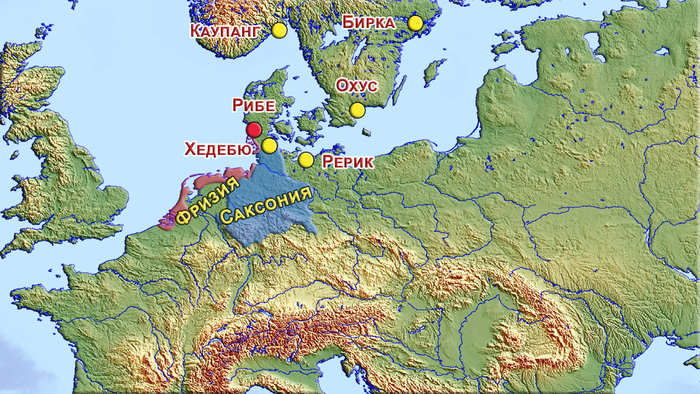
Жители этих поселений имели обширные региональные и межрегиональные контакты, благодаря как самостоятельным путешествиям, так и общению с торговцами, миссионерами и даже рабами.
Из вышеперечисленных, Рибе был самым ранним крупным торговым центром в Скандинавии, датируемый примерно 704-710 годом нашей эры. Он расположен недалеко от побережья юго-западной Ютландии, на территории современной Дании. На реконструированной карте видно, что основная территория поселения состояла из узких прямоугольных участков в два ряда по обе стороны улицы, идущей в направлении с СЗ на ЮВ, которую пересекала основная дорога регионального значения.
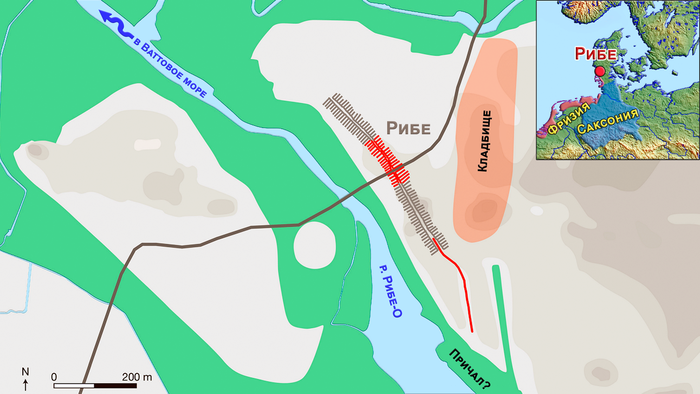
Восточнее поселения расположено кладбище, площадью около 8-9 га, которое послужило источником информации о культурной близости и географическом происхождении захороненных там людей.
Анализ соотношения стабильных изотопов стронция от 21 человека и изучение погребальных практик, позволяют по-новому взглянуть на раннее городское сообщество Рибе и намекают на сложные социальные и культурные процессы, связанные с передвижениями людей в контексте повышения роли городских поселений в Скандинавии.
Возникновение этих более или менее постоянных прибрежных поселений, чья экономика в большей степени сосредоточена на торговле и ремёслах, считается одним из наиболее важных социальных изменений в преимущественно земледельческих и скотоводческих сообществах Северной Европы в конце железного века и раннем средневековье.
И хотя есть много споров о том, что спровоцировало усиление роли городских центров в Скандинавии, особенно в контексте их связи с королевской властью, история формирования этих поселений эпохи викингов предполагает географическую мобильность и отмечена контактами между людьми с различными культурными связями и географическим происхождением, проживавшими как в непосредственной близости от городов, так и далеко за их пределами.
Торговый центр Рибе
Торговый центр Рибе, ещё с момента открытия археологами в 70-х годах XX века занимал особое место в археологии эпохи викингов и изучении раннесредневековой урбанизации. Сам торговый центр представляет собой плотно застроенный участок, датируемый VIII-X веками с богатой сборкой артефактов, указывающих на торговлю и специализированное производство ремесленных изделий. Останки домов, указывают на постоянное заселение эмпория. Однако также отмечены следы ещё одного, возможно сельскохозяйственного поселения, существовавшего почти в то же время, что и торговый центр с VIII-XI вв. Также на южном берегу реки Рибе с 2008 по 2012 год было раскопано христианское кладбище, основанное около 860 года.
В раннюю эпоху викингов торговая сеть Рибе расширилась, охватив территории современной Норвегии, побережье Балтийского моря, и в конечном итоге сформировала отдалённые связи со Средиземноморьем, а также с Ближним Востоком. Однако основная часть подтверждённых контактов Рибе указывает на южный регион Северного моря и долину реки Рейн, откуда в торговый центр поступали товары Восточно-Франкского королевства, такие как стекло, керамика и жернова.
Кладбище Рибе
Самое раннее задокументированное кладбище в Рибе расположено к востоку от торгового центра. На сегодняшний день около 100 археологических объектов было точно отнесено к нему, включая ряд маркеров могил, жертвоприношения и захоронения животных.
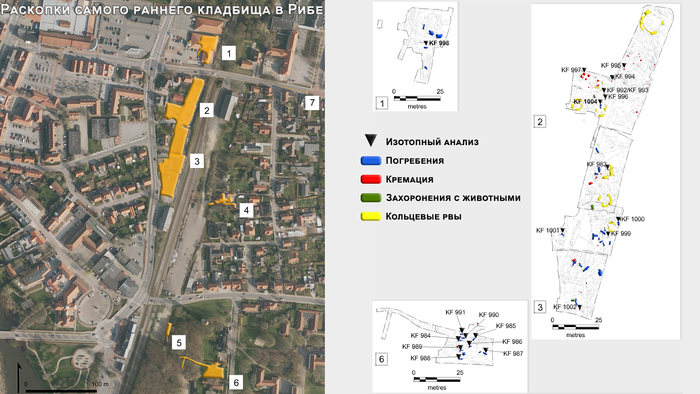
Основываясь на радиоуглеродном анализе артефактов и стратиграфии, кладбище была датирована VIII и началом IX века, а небольшое скопление могил в его южной части датируется IX и X веками. Стоит учитывать, что этот район практически непрерывно использовался с эпохи викингов до наших дней, поэтому некоторые следы могли не сохраниться. Помимо этого, распространённая в те времена практика кремации, подразумевает размещение отложений на поверхности или в неглубоких ямах, которые легко разрушаются, поэтому первоначальное количество могил эпохи викингов неизвестно. Предполагается, что их могло быть минимум несколько сотен.
Скорее всего, на кладбище были захоронены как жители торгового центра, так и жители сельского поселения. Но на нём, вероятно, хоронили и умерших гостей поселения.
Могилы Рибе, как правило, были плохо оснащены погребальным инвентарём. А из-за низкой сохранности несгоревших костей, скелетные останки сохранились не во всех захоронениях. В общей сложности, части скелетов людей и животных сохранились в 71-м из 100 захоронений, зафиксированных на кладбище. В некоторых случаях очертания скелета были задокументированы как более тёмная окраска песчаных отложений на дне могильной ямы, что позволяет добавить их к общему количеству определённых могил. В целом анализ выявил 67 человек, 57 из которых были захоронены в одиночных могилах, а 6 в парных. Оставшиеся 4 были связаны с нарушением контекста, что не позволило определить характер захоронения.
Анализ погребальных практик
Во времена викингов, Рибе находится на перекрёстке межрегиональных обменных маршрутов и в сложном культурном ландшафте. С одной стороны, его положение в Ютландии и к северу от системы датских укреплений или канала Даневирке, исторической границы Датского королевства, помещает Рибе в южно-скандинавскую культурную среду, а с другой стороны, поселение располагалось в непосредственной близости к Ваттовому морю, и в относительной доступности от других культурных групп, во Фризии и Саксонии, да и в регионе Северного моря в целом.
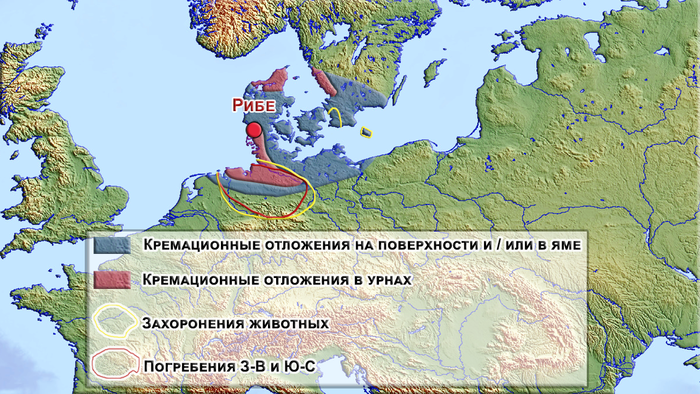
В определённой степени культурный ландшафт юго-западной Ютландии и фризских регионов имеет много общего с эмпорием, а погребальные практики в Рибе и окрестностях действительно отличаются от тех, которые засвидетельствованы для остальной части Ютландии с очень большим разнообразием методов захоронения. На кладбище присутствуют как погребения, так и более распространённая кремация.
Простые погребения
В случае с простыми погребениями, могилы отображают различия в ориентациях ям, положении тел, а также в наличии и отсутствии контейнеров для останков.
Могилы были ориентированы в основном с запада на восток или с юга на север, в большинстве случаев контейнеры не использовались, но есть несколько примеров гробов или лодок.
В могилах с запада на восток умершие, преимущественно, размещались в положении лёжа на спине, а в могилах с юга на север в полусогнутом положении.
Погребения редко содержали какие-либо захороненные предметы или животных, за исключением нескольких экземпляров одежды, украшений, монет и отдельных зубов лошади.
Кремации
Что касается кремаций, то это преимущественно небольшие, бесформенные круглые ямы, в которые помещали остатки костра с тем, что осталось от тел. Кремационные захоронения можно разделить на три основных типа:
1. Где останки костра были помещены в небольшую яму.
2. Где кремированные человеческие останки были осторожно извлечены из костра и помещены в урну.
3. Где остатки костра остались в виде небольших отложений на поверхности.
В качестве урн могла использоваться керамика как регионального типа, так и франкский импорт. В ямах и урнах человеческие останки часто смешивались с кремированными останками животных и различными предметами, такими как небольшие личные инструменты, снаряжение для верховой езды, украшения и т. д.
К животным относятся домашние млекопитающие, различные виды птиц и даже рыбы в одном из захоронений.
Иногда животных хоронили отдельно, о чем свидетельствует захоронение лошади со сбруей и седлом и захоронение собаки.
Теоретически различия в погребальных практиках могли быть обусловлены социальным статусом, возрастом и полом захороненных. Однако различия в степени затрат, связанных с процессом захоронения, как к примеру, количество и качество погребального инвентаря или использование бревенчатой лодки в качестве гроба, не могут считаться надёжными индикаторами социального статуса в конкретном контексте, поскольку каждый подтип захоронения включает небольшое число относительно более богатых захоронений, также как дети, взрослые, пожилые люди, а также мужчины и женщины были захоронены в различных из перечисленных типах захоронений. Таким образом, наиболее вероятным объяснением наблюдаемого разнообразия следует считать культурные связи, а не возраст на момент смерти или пол.
Погребальные практики и их распространение
Некоторые погребальные практики на самом раннем кладбище Рибе задокументированы как в других местах в непосредственной близости, так и за пределами описываемого региона. К примеру, кремированные останки одного взрослого мужчины (KF 994), в первой половине VIII века, были помещены в урну вместе с конской упряжью, а подобные случаи зафиксированы как в 15 км севернее Рибе, так и в Восточной Фризии или на севере современной Германии.
А кремационные отложения в урнах из керамики франкского типа со схожими артефактами, а иногда и видами животных, были распространены как в окрестностях Рибе и на Северо-Фризских островах VIII века, так и дальше по историческому побережью Фризии. Это относится к одному из детей (KF 999), которого кремировали, а останки поместили в кувшин, импортируемый из мест, где сейчас расположен современный район Майен-Кобленц Германии. Однако в то время кремационные захоронения не практиковались в восточной части христианского Франкского государства, где был изготовлен кувшин. Но повторное использование рейнских судов в качестве урн хорошо задокументировано вдоль побережья Фризии до Северо-Фризских островов.
А вот кремационные отложения в ямах были широко распространены по всей Южной Скандинавии и северной части Центральной Европы и не могут быть связаны с конкретным регионом.
Погребения с ориентацией могил с севера на юг, встречаются в Скании и на острове Борнхольм, подобные могилы, также отмечены вдоль побережья Фризии и на севере современной Германии. Погребальные ладьи с цельными бревенчатыми лодками или частями более крупных лодок задокументированы в Юго-Западной Ютландии от позднеримского железного века до эпохи викингов. Они также были широко распространены по всему региону Северного моря и особенно в Западной Скандинавии в эпоху викингов. Могилы, ориентированные с запада на восток, с гробами и без, как во Фризии, так и в Саксонии становились все более распространёнными в течение VIII века, вероятно, в результате христианского влияния, принесённого франкской экспансией, хотя они всё же были редки в Юго-Западной Ютландии вплоть до IX века. Захоронения лошадей и собак также встречаются в обоих регионах с очень близкими аналогами вдоль Эльбы. Однако стоит отметить, что выборки и сравнительные данные по указанному периоду Юго-Западной Ютландии, всё ещё ограничены, потому как погребальные практики в этом регионе хорошо задокументированы до 550 года, после чего захоронения почти исчезают из археологических записей вплоть до IX века, что возможно связано с кремациями на поверхности. Но для Фризии и Саксонии VIII века, погребальные практики хорошо задокументированы. А некоторые погребальные традиции, зафиксированные в Юго-Западной Ютландии, очень схожи с обычаями северного побережья Фризии, что указывает на тесные культурные связи между этими двумя регионами. Помимо этого, некоторые погребальные практики Рибе имеют параллели с регионом вдоль реки Эльба, но не имеют аналогов по Ютландии в целом. Как к примеру, в одном погребении кладбища Рибе, человек лежал в согнутом положении и головой на юг, что было редкостью для Рибе и Ютландии VIII и начала IX веков, однако подобные захоронения практиковались в регионе Эльбы в тот период.
Таким образом, с археологической точки зрения, самое раннее кладбище Рибе отражает погребальные практики не только Юго-Западной Ютландии, но и более отдалённых регионов.
В целом, учитывая дискуссии из предыдущих исследований, анализ практики захоронений, представленный в новой работе, позволяет предположить, что большая часть захороненных на самом раннем кладбище Рибе могла иметь, если не иностранное происхождение, то как минимум, крепкие культурные связи с людьми из регионов за пределами Юго-Западной Ютландии.
Результаты изотопного анализа
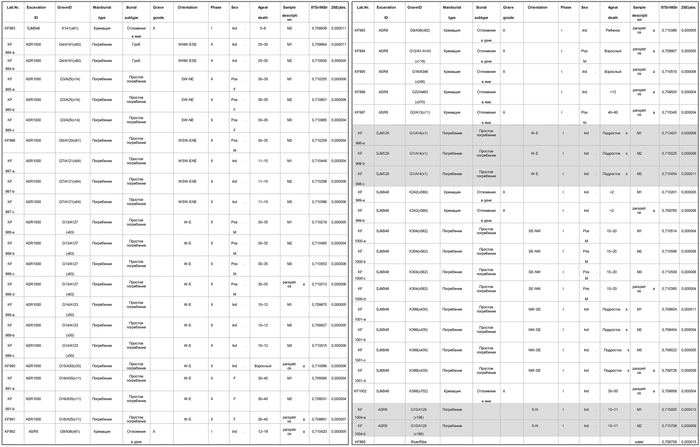
Несмотря на разницу в погребальных обрядах, только два человека из 21, показали значения соотношения изотопов стронция в пределах, указывающих на неместное происхождение. Однако есть и другие места со схожим диапазоном соотношения изотопов стронция за пределами современной Дании. Например, в настоящее время трудно различить людей родившихся и проживающих в Юго-Западной Ютландии и на побережье Фризии. Помимо этого, у одного из неместных представителей кладбища Рибе (KF 998), отмечена разница по соотношению изотопов стронция между первым и вторым моляром, что может указывать на переселение ещё в детстве из одного региона в другой. Его останки плохо сохранились и пол определить не удалось, но судя по зубам это был ребёнок. Он был похоронен в земле лёжа на спине, без гроба и головой на запад, без погребального инвентаря.
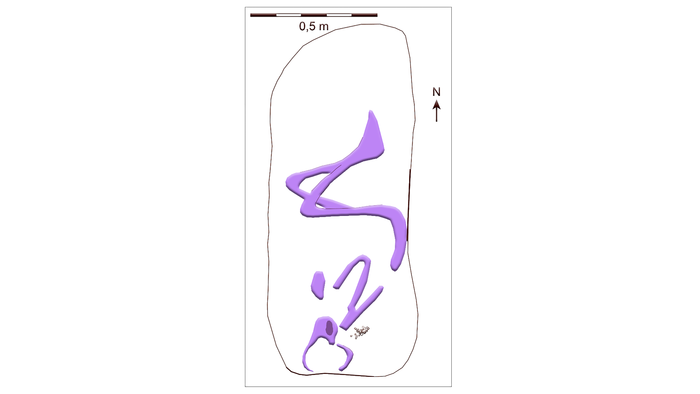
В данном случае, можно предположить христианское влияние, потому как подобные погребальные традиции отмечены в VIII и начале IX веков во франкских областях к югу, и чаще во Фризии и Саксонии. Принимая во внимание имеющиеся в настоящее время базовые знания о Северной и Центральной Европе в сочетании с археологическим контекстом, похоже, что этот маленький путешественник был родом из района Рейна в Центральной или Южной Германии.
А значения изотопов стронция у второго неместного представителя (KF 1004), указывают на то, что он прибыл в Рибе в более взрослом возрасте, около 10-11 лет и был погребён иначе чем вышеописанный представитель, а именно в согнутом положении головой к югу.
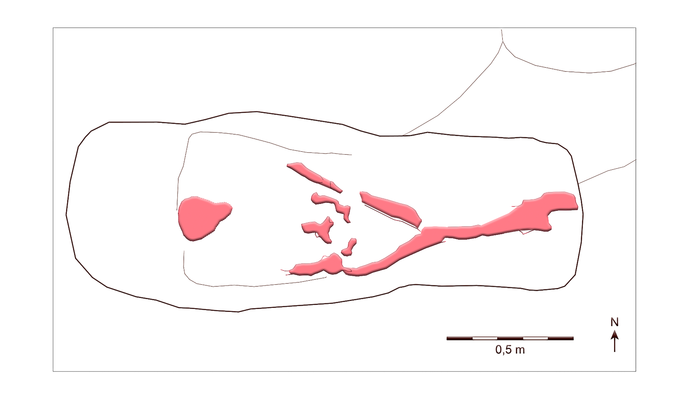
*В тексте статьи отмечено, что лицом на восток, но на иллюстрации на запад...
Тело укладывали на ткань или шкуры животных, точно определить не удалось. Несколько стеклянных бусин у затылка можно отнести к ожерелью или к украшению головного убора. Вполне возможно это было женское захоронение. Согласно датировке бусин, захоронение произошло между 760 и 800 гг. Головной убор со стеклянными бусами и ориентация тела, имеет близкую аналогию с погребением женщины на кладбище Рульсторф в земле Нижняя Саксония.
Помимо двух вышеописанных случаев, один взрослый человек из Рибе (KF985), возможно, женщина, демонстрирует небольшие различия между показателями стронция 1, 2 и 3 моляров, что может указывать на перемещение в детстве между территориями, у которых исходные диапазоны изотопов стронция попадают в диапазон для современной Дании. Захоронение представляет собой простое погребение без гроба и инвентаря, ориентированное с юго-запада на северо-восток, датируемое II фазой кладбища и не является признаком какого-либо регионального культурного сходства в Северо-Западной Европе того периода. Хотя набор данных слишком мал, тем не менее, выявленные случаи географической мобильности в подростковом возрасте заставляют задуматься, поскольку они предполагают, что взрослые торговцы, которые, как принято считать, перемещались по маршрутам сетей обмена, иногда сопровождались молодыми людьми.
Однако стоит отметить, что способы захоронения могут отражать представления людей, которые совершали обряды, а не ту культурную среду, в которой родился сам захороненный.
Сравнение с соседними регионами
Торговый центр Рибе обычно сравнивают с такими местами, как Бирка, Хедебю, Каупанг и Рерик, где были раскопаны большие кладбища, совмещённые с торговыми площадками.
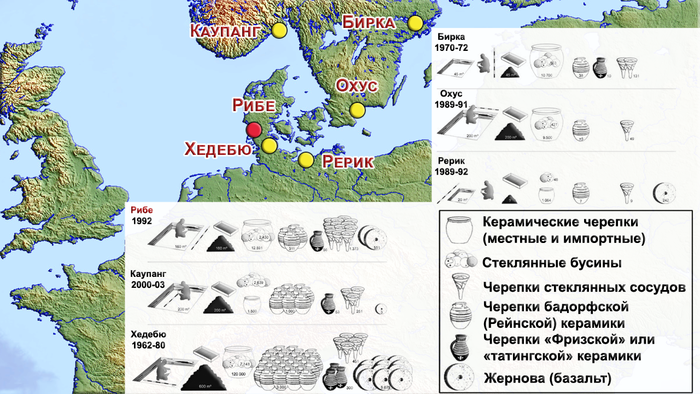
В шведском торговом центре викингов Бирке, к настоящему времени были проанализированы только 42 человека при помощи анализа изотопов стронция. Однако там также были выявлены как местных, так и неместных жители. В Рерике, на территории современной Германии (недалеко от Балтийского моря), значения изотопов стронция были измерены только для трёх человек, которые были интерпретированы как местные. Однако, как упоминалось ранее, эти значения также находятся в пределах базового диапазона для всего побережья Северного моря.
А вот в шведском городе Сигтуна, были проведены изотопные и генетические анализы 23 человек X-XII веков, которые с большей точностью указали на генетическое разнообразие и присутствие неместных представителей, а также выявили уже смешанных жителей. Но Сигтуна была основана более чем через два столетия после Рибе, в совершенно ином социальном, культурном, политическом и экономическом контексте, что не позволяет проводить прямое сравнение.
Выводы
В контексте морских торговых сетей в раннесредневековой Северо-Западной Европе поток товаров предполагает существование связующих общин из различных регионов. Хотя считается, что географическая мобильность человека является предпосылкой для таких обменов, социальная динамика, характеризующая торговые поселения – эмпории, через которые эти сети были созданы и степень, в которой они принимали мобильные группы и более постоянно оседлые сообщества, все ещё нуждается в дальнейшем изучении. Результаты анализа останков с самого раннего кладбища Рибе показывают, что большинство проанализированных людей имеют значения изотопов стронция, которые попадают в ранее охарактеризованный базовый диапазон современной Дании, включая и Юго-Западную Ютландию, где находится Рибе. С другой стороны, разнообразие погребальных практик отражает культурное сходство, которое может быть связано с общинами за пределами современной Дании, например, с побережьем Фризии и северной частью региона реки Эльбы в Германии. Эти районы имеют похожие значения изотопов стронция, которые частично совпадают с Данией, что ограничивает возможность определения движения между этими регионами без дополнительных методик. Таким образом, основываясь на сочетании данных археологии и соотношений изотопов стронция, нельзя исключить возможности того, что некоторые из людей, которые были отнесены к местным жителям Рибе, происходят из регионов со схожими значениями изотопов стронция. И несмотря на расстояния, контакты между этими регионами могли осуществляться вдоль берегов Ваттового моря.
Только два человека из 21 имели значения изотопов стронция, которые указали на неместное происхождение захороненных, при этом они были детьми. Поэтому в свете современных знаний о поселении Рибе эпохи викингов, новые результаты кажутся несколько удивительными. Хотя материальная культура, обнаруженная в жилищах и мастерских торгового центра, предполагает наличие или, по крайней мере, контакт с людьми, принадлежащим к общинам из местных, региональных и отдалённых районов, новое исследование не выявило масштабных путешествий из отдалённых мест. Но это можно списать на размер выборки, а в Рибе на самом деле проживало и захоронено больше неместных жителей. К тому же, неместные дети, выявленные в исследовании, вряд ли путешествовали в одиночку.
А в целом результаты показывают, что если торговый центр Рибе действительно привлекал людей, которые родились за его пределами для переселения и торговли, то они могли происходить из районов Ваттового моря и Эльбы, которые демонстрируют высокую степень культурной интеграции с Юго-Западной Ютландией и схожие значения изотопов стронция.
Источник: Croix S, Frei KM, Sindbæk SM, Søvsø M (2020) Individual geographic mobility in a Viking-Age emporium—Burial practices and strontium isotope analyses of Ribe’s earliest inhabitants. PLoS ONE 15(8): e0237850. doi.org/10.1371/journal.pone.0237850
Прежние исторические свидетельства о том, как гиксосы или «правители чужих земель», захватили власть и образовали XV династию фараонов Древнего Египта (около 1640-1530 гг. до н. э.) весьма сомнительны. Древнеегипетский историк и жрец Манефон на протяжении веков был единственным рассказчиком об их взлёте, правлении и падении. Однако сам Манефон жил более чем через тысячу лет, после династии гиксосов.

Он описывал родоначальников XV династии, как предводителей армии захватчиков, вторгшихся с северо-востока и захвативших северо-восточную дельту Нила во время Второго переходного периода в XVII—XVI веках до н. э.
При этом сами рассказы Манефона сохранились только в трудах более поздних историков, таких как Иосиф Флавий, и, несмотря на свою предвзятость и ненадежность, это были единственные известные источники о гиксосах на протяжении веков. Даже после расшифровки иероглифов, источники с описанием жизни гиксосов оставались скудными и ненадежными, преимущественно из-за древней цензуры и пропаганды. Где гиксосы ассоциировались с беспорядками, хаосом, а также с ритуальным насилием для поддержания порядка и легитимности власти. Правление гиксосов продолжало осуждаться фараонами Нового царства, такими как Хатшепсут, к примеру.
В более ранних исследованиях правителей гиксосов объединяли в целую этническую группу, что ещё больше осложнило понимание их происхождения. Стоит отметить, что в современной науке связь гиксосов с исходом евреев из Египта больше не является центральным предметом научных исследований, хотя некоторые учёные продолжают утверждать, что повествование об Исходе могло развиться из коллективных воспоминаний об изгнании гиксосов из Египта.
За время раскопок в древней столице гиксосов Аварисе особенно в последние годы, появилась возможность исследовать обстоятельства, при которых гиксосы пришли к власти.
Последние десятилетия исследований дали свидетельства, ясно указывающие на ближневосточное происхождение правящего класса, известного как гиксосы. На это указывают неегипетские черты керамики, погребальных обрядов, украшений, оружия и даже внутренней и культовой архитектуры.

Хотя это была не иностранная элита, прибывшая непосредственно из чужих земель, как рассказывал Манефон, а люди неегипетского происхождения, которые родились и выросли в дельте Нила.
Археологические свидетельства также не подтверждают рассказ Манефона о том, что гиксосы возглавляли армию захватчиков, вторгшихся с северо-востока, вместо этого предполагается, что те, кто стал правителями гиксосов, имели западноазиатское происхождение, но жили в Египте в течение многих поколений.
Разнообразие новых данных и находок позволяет провести прямое сравнение культуры гиксосов с Левантом и более обширными территориями Ближнего Востока, хотя до настоящего времени не было раскопано ни одной гробницы, которая принадлежала бы правителю гиксосов.
Участок раскопок Телль эль-Даба
Изучение захороненных на кладбищах Телль эль-Даба, на юге Авариса, дает возможность непосредственно оценить происхождение этих людей и оценить вопросы, касающиеся сроков и механизмов прихода гиксосов к власти.

Тем более, что археология показывает постоянное присутствие выходцев из Западной Азии в Аварисе на протяжении более 150 лет ещё до начала правления гиксосов.
На участке Телль эль-Даба, расположенном в Северо-Восточной дельте Нила, выявлена стратиграфическая последовательность, протяженностью более 500 лет. Это поселение, основанное в XII династии, с XIII династии было известно, как Хутварет. Во времена Среднего царства этот город был административным центром и портовым городом, который набирал силу, чтобы в конце концов стать столицей регионального царства гиксосов.

Будучи известным как Аварис, портовый город принимал представителей различных народов как ближнего, так и дальнего зарубежья, которые делились своими взглядами на жизнь и обсуждали технологии, религию, а также политику. А после того как гиксосы потерпели поражение от XVII династии фараонов, параллельно правивших в Фивах, Аварис был в значительной степени заброшен, примерно, после 1550 года до н. э.
Кстати, гиксосам приписывают некоторые технологические новшества, такие как лошадь и колесница, серповидные мечи – хопеши, а также композитные луки, но эти предположения оспариваются. Хотя металлические изделия в археологических слоях на участках раскопок A/I и A/II в Телль эль-Даба появляются после слоя E/1 и D/3 в течение периода правления гиксосов.
Участок Телль эль-Даба примечателен тем, что на кладбищах в его пределах захоронены люди, жившие до и во время правления гиксосов, поэтому в новой работе рассматривается 3 главных вопроса, связанных с притоком и перемещением людей, похороненных в Телль эль-Даба:
1. Есть ли хронологическая закономерность в перемещении людей? А именно, был ли приток в Телль эль-Даба больше во времена XII или XV династии?
2. Кого было больше среди неместного населения, мужчин или женщин?
3. Можно ли с уверенностью определить происхождение неместного населения?
Первые два вопроса касаются времени появления гиксосов и механизмов захвата власти. Ведь приток людей, особенно мужчин из-за пределов Египта к началу правления гиксосов придал бы большую вероятность вторжению. И наоборот, более-менее равное количество мужчин и женщин, могло бы свидетельствовать о переселении в дельту Нила семей в связи с её экономической привлекательностью.
А вот ответ на третий вопрос помог бы разгадать самую главную загадку XV династии и раскрыть места откуда собственно и пришли гиксосы.
Результаты
Не вдаваясь в подробности методик, при сопоставлении с местными значениями стронция более половины исследуемых людей из Телль эль-Даба провели детство за пределами дельты Нила, независимо от участка раскопок.

Также отмечены достоверные различия в значениях стабильных изотопов стронция у людей, живших до прихода гиксосов к власти и во времена их правления. При этом большое количество мигрантов отмечено в периоды до гиксосов, причём женщин среди мигрантов было больше, что явно не вписывается в модель завоевания.
Выводы
Нил был основным источником воды для питья и полива сельскохозяйственных культур, поэтому значения стронция для большинства людей живших в Древнем Египте, были бы близки к местному диапазону Телль эль-Даба. При этом люди, выросшие вдоль Нила, вероятно, демонстрируют слишком близкие значения, чтобы их различать, например, в Верхнем и Нижнем Египте. Поэтому стоит учитывать, что неместные жители исследуемого региона, жившие к югу от дельты Нила, показали бы те же значения стронция, что и местные жители. Т.е. среди проанализированных людей, которые показали значения соотношения изотопов стронция как у местных жителей, могли быть выходцами из Верхнего Египта, а именно Мемфиса, Фив или даже из Нубии южнее. И они не могут быть идентифицированы с помощью анализа изотопов стронция.
Большое количество мигрантов до династии гиксосов в сочетании с предыдущими археологическими данными, поддерживают выводы о том, что гиксосы пришли к власти без вторжения в верхнюю часть дельты Нила, они уже там проживали и были знакомы с местной системой власти. Кроме того, большое количество неместных женщин также не стыкуется с вторжением армии. Такое соотношение мужчин и женщин характерно для семей, переезжающих в более благополучный с экономической точки зрения регион. Но довольно высокая доля женщин около 77%, требует более тщательного рассмотрения.

Предполагается, что раскопанные кладбища и захоронения в большей степени представляют элиты города, чем рядовых жителей, и вполне возможно, что эти женщины прибыли в регион для заключения браков, скрепляющих союзы с могущественными семьями за пределами Нила. Во времена Среднего царства и Второго переходного периода существует много свидетельств того, что мужчины с египетскими именами брали в жены женщин с неегипетскими именами и такое отношение к браку сохранилось и в XVIII династии. Неместные женщины могли вступать в брак с представителями высокопоставленных египетских семей, при этом браки египетских женщин с высокопоставленными иностранцами не практиковались.
Также немаловажным результатом этого исследования было и то, что широкий диапазон значений у жителей Телль эль-Даба, говорит о том, что неместные жители, как до, так и во время правления гиксосов, происходили не из одного конкретного географического места, а из широкого разнообразия источников в Западной Азии.

Хотя правители XV династии фараонов могли иметь общее и даже родственное происхождение, дельта Нила в те времена, да и задолго до правления гиксосов представляла собой многокультурный центр.
В целом, исследователи не ставят под сомнение левантийское происхождение правителей гиксосов из-за их имен, материальной культуры и архитектуры, новые данные бросают серьёзный вызов классическому повествованию о гиксосах как о завоевателях. Исследование также подтверждает теорию о том, что элита гиксосов была не из единого места происхождения, а из обширных мест Западной Азии, чьи предки переселились в Египет во время Среднего царства и жили там в течение веков, а затем пришли к власти.
Источник: Stantis C, Kharobi A, Maaranen N, Nowell GM, Bietak M, Prell S, et al. (2020) Who were the Hyksos? Challenging traditional narratives using strontium isotope (87Sr/86Sr) analysis of human remains from ancient Egypt. PLoS ONE 15(7): e0235414. doi.org/10.1371/journal.pone.0235414
Примечание
Предлагаю вашему вниманию более подробный обзор оригинала научной работы, которая уже упоминалась на pikabu под названием "Анализ ДНК подтвердил теорию Тура Хейердала". Но выводы авторов статьи, по поводу соответствия полученных результатов предположениям Тура Хейердала, читатели могут оценить самостоятельно, прочитав этот обзор. Особенно касательно заселения острова Пасхи.
Вероятность контактов между древними полинезийцами и коренными американцами давно интересовала исследователей. Сторонники таких контактов опирались на батат и бутылочную тыкву среди археологических находок в Полинезии и доколумбовой Америке, но нигде больше. Однако критики утверждали, что распространение этих культур могло происходить не только при помощи человека. Норвежский исследователь Тур Хейердал спорно предположил, что доисторическое население Южной Америки сыграло важную роль в заселении Восточной Полинезии и особенно острова Пасхи (Рапануи). Несколько ограниченных молекулярно-генетических исследований пришли к противоположным выводам, и эта возможность так же горячо оспариваться по сей день. Рапануи считался наиболее вероятным местом контакта с древними американцами. Поэтому предыдущие исследования, были сосредоточены на этом месте, как самом близком к Америке населённом острове Полинезии с наиболее развитой мегалитической культурой. В новой работе были проанализированы геномы 807 человек из 17 островных популяций Полинезии для выявления признаков примеси с коренными американцами из 15 групп Тихоокеанского побережья и континентальными связанными популяциями.
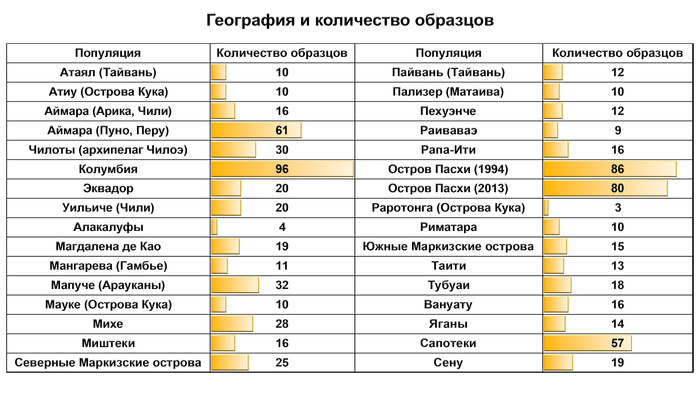
Множественные события смешения в Полинезии
Различные методы и анализы показали, что тихоокеанские островные популяции характеризуются значительным полинезийским компонентом, но у многих также присутствует европейский компонент от колониальной примеси. Однако, в Восточной Полинезии была обнаружена примесь коренных американцев, даже при использовании образцов доколумбового периода.
При этом на самых восточных Полинезийских островах, в частности островах Пализер, Маркизских островах, Мангарева и острове Пасхи, выявлено два компонента предков коренных американцев, в отличие от других. Эти компоненты характерны как для современных, так и древних популяций Центральной и Южной Америк. При этом южноамериканский компонент, который наиболее распространён у коренных народов Америки мапуче и пехуэнче из Чили, у представителей Рапануи, увеличивается пропорционально европейскому компоненту. Что согласуется с идеей о том, что индейский компонент прибыл на остров Пасхи вместе с испанским европейским компонентом благодаря миграции смешанных чилийских представителей после того, как остров был аннексирован Чили в 1888 году.
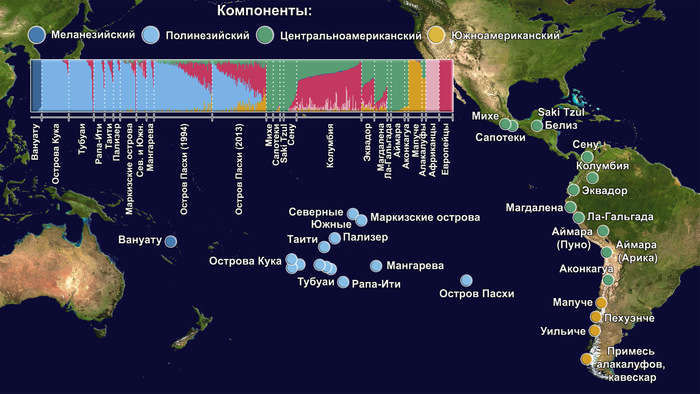
В отличие от этого, центральноамериканский компонент, характерный для коренных мексиканцев (михе и сапотеков) и коренных колумбийцев (сину), на Рапануи ассоциируется только с полинезийским компонентом, но никак не с европейским или южноамериканским компонентом, а это говорит о том, что центральноамериканский компонент прибыл на остров Пасхи независимо от европейского компонента. Кроме того, в отличие от южноамериканского, центральноамериканский компонент коренных американцев слабо отличается между представителями Рапануи, а это указывает на то, что он происходит от более древнего события примеси. Сегменты ДНК коренных американцев у представителей острова Пасхи имеют совокупное распределение длины, которое указывает на первый контакт за несколько столетий до того, как в этом регионе Тихого океана появились европейцы.
Примечательно, что центральноамериканский компонент встречается и на других отдаленных к востоку полинезийских островах, таких как Пализер, Маркизские острова и остров Мангарева, и имеет столь же раннюю расчётную дату примеси.
Помимо этого, европейское происхождение у полинезийцев соответствует европейским популяциям, которые колонизировали острова. Например, европейский компонент у жителей Французской Полинезии соответствует французским образцам, а жители острова Пасхи с южноамериканской примесью смещены в сторону испанских эталонных образцов, однако другие представители острова имеющие европейскую примесь, но без южноамериканской, объединяются в основном с французскими образцами, что согласуется с французским происхождением первых европейских поселенцев на Рапануи.
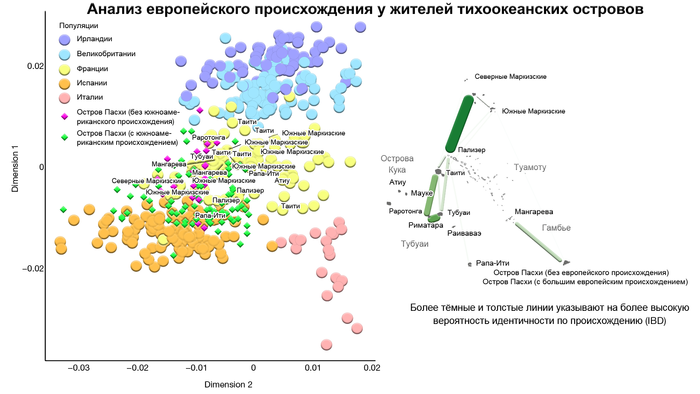
Используя метод идентичности по происхождению для европейских сегментов ДНК у полинезийцев, исследователи обнаружили, что многие жители островов Французской Полинезии по европейскому компоненту группируются вместе, при этом отдельно от жителей острова Пасхи. Единственная европейская связь Рапануи с островом Мангарева может отражать перемещение французских католических миссионеров от острова Пасхи в 1871 году.
Родословная коренных американцев в Полинезии
В соответствии анализом примесей, который показал наличие центральноамериканского компонента у островитян Тихого океана, все эти предки попадают в состав или рядом с индейским народом Колумбии — сину. Исключение составляют только представители Рапануи с высокой долей европейского происхождения. Как и ожидалось, их компонент коренных американцев близкий к мапуче и пехуэнче, вероятно, попал на остров параллельно с европейским компонентом благодаря уже смешанным чилийским переселенцам. А вот жители острова Пасхи без европейского происхождения также группируются с колумбийским народом сину.
Помимо чилийской аннексии острова Пасхи в 1888 году и редких случаев взаимодействий с экипажами кораблей, единственными зарегистрированными событиями, потенциально связывающими жителей тихоокеанских островов с коренными американцами, являются набеги перуанских работорговцев в 1862-1863 годах. В течение этого времени тысячи жителей тихоокеанских островов были насильно вывезены в Перу в качестве подневольных рабочих, в том числе 1407 жителей Рапануи. Однако после международного скандала было организовано несколько рейсов по возвращению островитян домой, но вспышки оспы на борту привели к тому, что лишь малая часть пассажиров вернулась в Полинезию живой. В наборе данных из этого исследования было зарегистрировано 24 вернувшихся человека, только на двух островах – Рапануи и Рапа-Ити.
Однако учитывая, что в Полинезию вернулось очень мало людей и все они идентифицировали себя как островитяне, а их плен в Перу длился всего несколько месяцев, маловероятно, что этот эпизод привел к какой-либо существенной примеси родословной коренных американцев в полинезийцев. Но такие объяснения имели место быть, ведь для примеси достаточно и нескольких минут. Хотя в любом случае, компонент коренных американцев, который прослеживается у самых восточных островитян, в том числе на отдаленных островах, не затронутых возвращением, происходит от коренного американского населения, которое проживало к северу от Перу.
Локализация предков коренных американцев, которые внесли свой вклад в полинезийцев, согласуется с несколькими лингвистическими, историческими и географическими наблюдениями, которые подтверждают их происхождение в северной части Южной Америки.
Хотя поверхностное сходство между статуями на отдаленных восточных Полинезийских островах и статуями доколумбового периода в окрестностях Сан-Агустина Колумбии уже давно замечено, более весомые аргументы опирались на полинезийское слово, обозначающее батат – "кумара". Похожее слово было связано с северной частью Южной Америки и Перу, где оно и возникло. Помимо этого, именно к северу от Перу природный ландшафт меняется от пустыни к лесам, пригодным для строительства лодок. Авторы предполагают, что именно от тихоокеанского побережья Колумбии и Эквадора американские индейцы отправились в путь для торговли с Мезоамерикой на больших парусных плотах, сделанных из бальзового дерева в период с 600 по 1200 год.
Моделирование ветров и течений с тихоокеанского побережья Северной и Южной Америки показало, что дрейфующие плоты, отплывающие из Колумбии и Эквадора, с наибольшей вероятностью достигли бы южных Маркизских островов Полинезии, за которыми следует группа островов Туамоту. Оба этих архипелага лежат в самом сердце региона, где был обнаружен компонент коренных американцев из Колумбии.
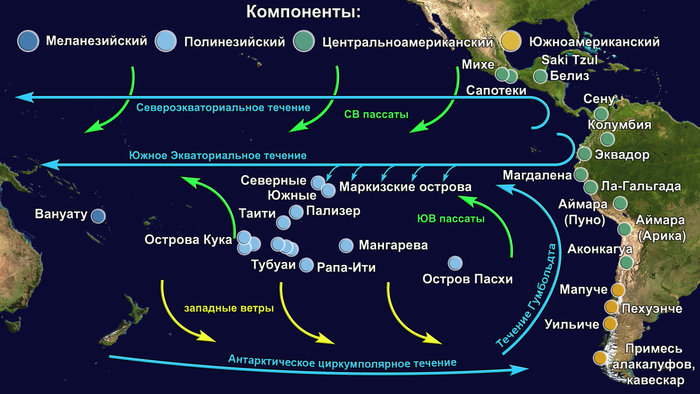
Круглогодичные ветры пассаты и Южное Экваториальное течение движутся в этих широтах с востока на Запад, направляя лодки из северной части Южной Америки к архипелагам. Тур Хейердал на плоту из бальсовых брёвен Кон-Тики испытал на себе эти природные явления в своём знаменитом путешествии из Южной Америки в Полинезию. По той же причине Маркизские острова и Туамоту были бы наиболее вероятным начальным этапам для путешествия из Полинезии В Америку с подветренной стороны.
Датировка контактов между полинезийцами и коренными американцами
Предполагаемые даты примеси между коренными американцами и полинезийцами, схожи на разных островах, за исключением острова Пасхи, где рапануйцы, не имеющие европейского (колониального) происхождения, показывают несколько более позднюю дату примеси от коренных американцев – 1380 год, в отличие от Маркизских островов, Мангарева и Пализер, где эта примесь зафиксирована от 1150 до 1230 года. И скорее всего это было одиночное событие примеси.
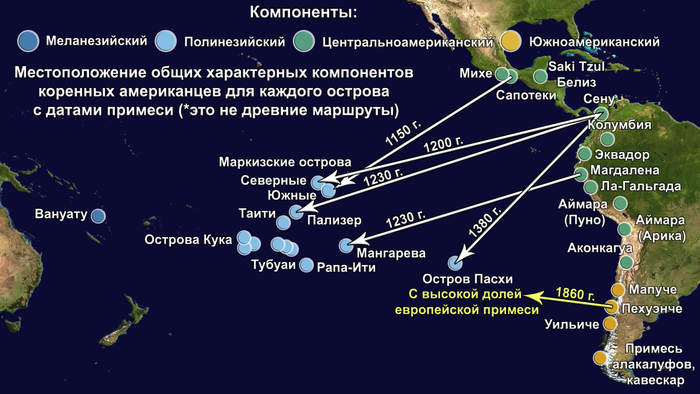
Несмотря на примечание на графике, многие читатели воспринимают стрелки, как маршруты миграций, но это не так.
При этом стоит отметить, что остров Пасхи является одним из самых сложных мест для изучения и датировки доисторических контактов с коренными американцами в Восточной Полинезии.
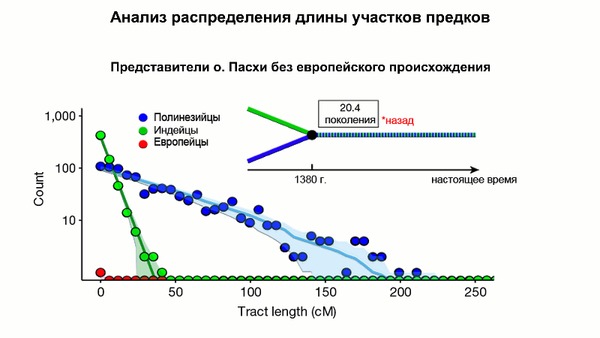
А вот европейская примесь хорошо соответствует периоду европейской колонизации в регионе, через сотни лет после контактов полинезийцев и коренных американцев.
Итоги
В своей работе авторы приводят довольно убедительные доказательства древнего контакта полинезийцев и коренных американцев, наиболее тесно связанных с древними жителями современной Колумбии, за сотни лет до европейской колонизации Америки и островов. Более ранняя дата примеси, около 1150 г., получена для самого южного острова Маркизских островов – Фату-Хива.

Фату-Хива
И эта примесь скорее произошла, ещё до заселения острова Пасхи, на который этот компонент попал уже с заселившими его людьми. Помимо этого, наличие примеси не обязательно указывает на целенаправленный маршрут индейцев, более чем за 6 тысяч километров. Скорее всего случай, течения и ветры сыграли в этом деле ключевую роль. Хотя время контакта совпадает с периодом, когда, по данным некоторых исследований полинезийцы совершали самые дальние путешествия между островами Тихого океана, от Гавайев до Новой Зеландии и Рапануи.
Поэтому исследователи не исключают и альтернативные сценарии. По одному из маловероятных, но не исключённых сценариев, полинезийцы доплыли до Америки и вернулись назад уже с индейцами или в составе смешанной популяции. А по-другому, когда полинезийцы прибыли на Маркизские острова, там уже проживали коренные американцы – "занесённые ветром", образно говоря. Потому как даты примеси совпадают с датами заселения островов.
Кстати, обратите внимание, что результаты исследования отличаются от предположений Тура Хейердала.
А в целом результаты работы демонстрируют важность генетических исследований современных популяций, которые позволяют при больших размерах выборки и в сочетании с другими методами, разгадывать сложные исторические загадки. Это уже не косвенные и противоречивые лингвистические, ботанические и археологические данные.
Источник: Ioannidis, A.G., Blanco-Portillo, J., Sandoval, K. et al. Native American gene flow into Polynesia predating Easter Island settlement. Nature 583, 572–577 (2020). doi.org/10.1038/s41586-020-2487-2
Цыгане — это преимущественно кочующая индоарийская этническая группа, без собственной письменной истории, однако за 15 веков распространившаяся, по обширным территориям. Наибольшее их количество сосредоточено в регионах Восточной и Центральной Европы, но они в большом количестве живут на Пиренейском полуострове, а также на Кавказе, Ближнем Востоке и в Америке. Точное количество цыган трудно оценить из-за отсутствия надежных методов переписи, поэтому в разных источниках данные варьируются от 4 до 15 млн. человек. Сразу отмечу, что у описываемых групп людей есть различные самоназвания, да и разные народы их также называли по-разному. Также акцентирую внимание на том, что речь в данном видео преимущественно о европейских популяциях цыган. Почти не касаясь цыган – дом, лом, люли и других.
Сравнительная лингвистика помещает родину европейских цыган в Индии, особенно в северо-западном регионе, поскольку цыганский язык тесно связан с пенджабским и кашмирским. Однако социальная организация и культурная динамика в индийских популяциях такова, что люди, проживающие в тесно связанных языковых группах и даже в одном и том же регионе, имеют разные пропорции генетических компонентов, связанных с предками южных и северных индийцев. Что не позволяет рассматривать их как генетически однородные группы и затрудняет поиск происхождения цыган, основанный исключительно на лингвистических данных.
Индийская генетическая составляющая цыганской популяции была впервые предложена после выявления общих болезнетворных мутаций у индийских и пакистанских пациентов. Анализ однополых маркеров позволил установить индийское происхождение некоторых материнских и отцовских линий, обнаруженных у цыган. Это относится к линиям М5, М18, М25, М35, митохондриальной гаплогруппы M и к линии М69 гаплогруппы Y-хромосомы Н.
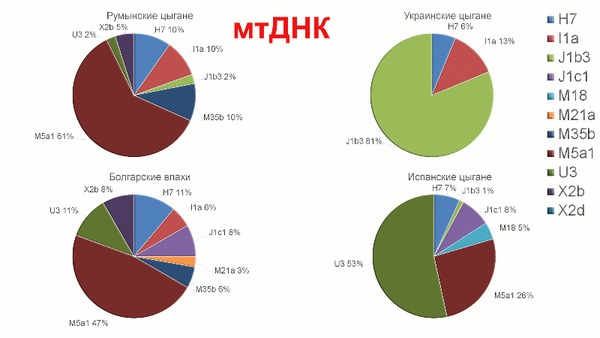
Кроме того, общегеномные исследования показывают, что европейские цыгане происходят от уменьшенного числа популяции-основателей или протоцыган, прародиной которых был нынешний штат Пенджаб на северо-западе Индии.
Согласно предыдущим историческим и антропологическим свидетельствам, предковые популяции цыган мигрировали из Северо-Западной Индии на Балканский полуостров, через Персию, Армению и Малоазиатское нагорье, примерно в IX-X веке нашей эры. А в XI-XII веке некоторые цыгане расселились по Балканскому полуострову, образовав соответственно балканских цыган. Другие группы распространились по Дунайским княжествам (современная Румыния, Молдова и Венгрия), где их принудили к рабству, так получились цыгане-влахи.
Под влахами (Vlax Roma) в данном случае подразумеваются группы цыган, говорящие на влашских диалектах цыганского языка
А группы будущих карпатских цыган или ромунгров, рассеялись по всей Австро-Венгерской империи. И наконец, другие небольшие группы переселились в Северную, Центральную и Западную Европу (северо-западные цыгане). Прибытие цыган на Пиренейский полуостров в начале XV века, подтверждается рядом иберийских исторических записей, упоминающих о присутствии цыганских групп в Сарагосе и Барселоне в 1425 и 1447 годах соответственно.

Выше описана довольно упрощённая модель, а в целом распространение цыган по Ближнему Востоку, Кавказу и Европе было очень сложным процессом, в ходе которого цыганские популяции получали существенные потоки генов из различных европейских, ближневосточных и кавказских, нецыганских популяций. Помимо этого, обмен генами происходил и между генетически отличающимися популяциями цыган. Общегеномные данные показали, что у цыган до 80% западноевразийских предков (с учётом популяций Ближнего Востока и Кавказа), в то время как остальные 20% происходят из южноазиатских источников. Однако здесь стоит учитывать, что часть западноевразийского компонента, до 15%, уже присутствовала в Южной Азии задолго до формирования прото-цыганских популяций, ещё около 4 тыс. лет назад в виде предков северных индийцев. Более подробно описано в ролике: Популяционная история Центральной и Южной Азии по данным древней ДНК.
Также хочу обратить внимание зрителей, на то, как генетически изменялись популяции цыган всего за полторы тысячи лет, несмотря на практики заключения браков в пределах своей группы. Ведь в итоге цыгане состоят из разнородной и субструктурированной мозаики популяций, которые демонстрируют культурно-исторические и лингвистические различия не только с соседними популяциями, но и между собой. Это может быть хорошим примером, для некоторых людей, представляющих популяции с эндогамными социальными практиками, как что-то единое и неизменное в генетическом плане на протяжении тысяч лет. Это любителям великих предков на заметку.
При этом эндогамия приводит к различному спектру наследственных заболеваний, что и отмечено у цыган, по сравнению с другими популяциями. Поскольку географически рассредоточенное кочевое население было социально изолировано, а также часто подвергалось преследованиям начиная со Средневековья и до наших дней.
Исторические записи подтверждают преследование и социальную маргинализацию, которой подвергались популяции цыган с момента прибытия в Европу. Также предпринимались попытки их насильственной ассимиляции, например, в Габсбургской монархии или Норвегии с XVIII по XX века, а также в СССР.
Структура популяции цыган
Образцы цыган из разных исследований, на графике анализа главных компонент расположились между европейцами и популяциями Южной Азии, в соответствии с их южноазиатским происхождением и последующем смешением с европейцами.
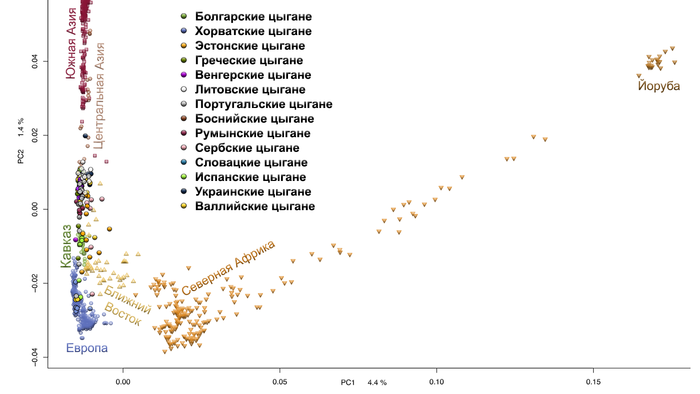
Анализ примесей показал два основных компонента предков: западноевразийский и южноазиатский, что еще раз подтверждает южноазиатское происхождение цыган и недавнюю европейскую примесь.

А при сравнении цыганских групп между собой, исследователи обнаружили четкое разделение между северными и западными группами, испанскими и прибалтийскими цыганами, а также между северо-западными цыганами и другими группами.
Также не обошлось и без сюрпризов для добровольцев, у одного представителя влахов из Венгрии вообще не обнаружилось цыганской родословной, после чего этот образец ДНК был удален из последующих анализов.
Кстати, вопрос: кем считать этого человека, если он сам считает себя цыганом?
Формирование цыганских популяций и источники примеси
После того, как популяции прото-цыган покинули территорию Индию были зафиксированы события примеси с европейскими популяциями около 1270–1580 гг. Этот интервал дат вписывается в периоды, когда появляются первые исторические записи о цыган в странах Европы. Более ранние даты примеси из Балканского полуострова и Центральной Европы, они указывают на то, что расселение цыган по Европе происходило через Балканы. В результате примеси с популяциями по пути следования, у цыган от южноазиатского источника осталось только около 35%. При этом по распределению аллелей этот источник схож с пенджабцами из северо-западной Индии и не только, так как пенджабцы генетически неоднородны и не группируются вместе в связи со сложной и неоднородной структурой населения в Индии с различными пропорциями предков южных и северных индийцев. Но большая часть южноазиатского происхождения цыган в основном принадлежит группе лиц из Пенджаба с меньшим западноевразийским компонентом. Также отмечен вклад и от популяций говорящие на дравидийских языках, как народ ирула, к примеру, представители которого также демонстрируют низкие уровни западноевразийского происхождения.
Стоит отметить, что большая схожесть цыган с современными популяциями Пакистана, чем с Северо-Западной Индией, обусловлена потоком европейских генов на эту территорию уже после того, как прото-цыгане покинули Индию. Это также стоит учитывать.

В целом, эти данные позволяют предположить, что наиболее вероятным представителем южноазиатского происхождения прото-цыган является наследственный источник, описанный как смесь современных южно-азиатских групп с низким западно-евразийским вкладом. Анализ идентичности по происхождению, также указал на Пенджаб как предполагаемый регион происхождения предков цыган в пределах Индийского субконтинента. При этом формирование цыганского населения, происходило как единое событие.

А среди популяций, которые внесли наибольший свой вклад в генофонд цыган по ходу их движения в сторону Европы, были армяне, а затем турки. При этом, после прибытия цыган в Европу, дальнейших примесей с представителями Ближнего Востока и Кавказа не наблюдалось.
Что касается европейского вклада, то нет никаких свидетельств того, что какие-то отдельные популяции являются источником потока генов в конкретную группу цыган, хотя юго-западные и юго-восточные европейцы, по-видимому, вносят больший вклад. В цыганских группах Балкан и Центральной Европы европейская примесь составляет около 60%, и увеличивается до 80% по мере удаления от Балканского полуострова. Северные и западные цыгане, включают наибольшее количество европейского происхождения. У цыган, мигрировавших в Северную и Юго-Западную Европу, балканский компонент оставил отличительный след, хотя он был сильно переконфигурирован примесью в Балтийском регионе и на Пиренейском полуострове. Демографическая модель предполагает несколько событий генетических расколов в цыганских популяциях. После отделения их от индийских популяций около 1600-2000 лет назад, первый раскол произошел уже на Балканском полуострове между предками балканских цыган и влахов, и группами мигрантов за пределами Балкан (общими предками карпатских и северо-западных цыган). Два более поздних раскола произошли между карпатскими и северо-западными цыганами, а также между балканскими цыганами и влахами.
Изменение эффективного размера популяции цыган на протяжении поколений
Анализируя все группы цыган вместе, исследователи обнаружили, что эффективный размер предковой популяции цыган перекрывался с таковым у северных индийцев, примерно, до 125 поколений назад, в соответствии с происхождением цыган в этом регионе. Однако в последующие годы эффективный размер популяции предков цыган постоянно сокращался и начал отличаться от эффективного размера популяции Северной Индии. При этом группы северных и южных индийцев не продемонстрировали столь резкого сокращения численности населения. Расчёт показал, что минимальный эффективный размер популяции цыган между 1159 и 1333 годом н. э., составлял 871 - 1160 человек, после чего эффективный размер немного увеличился и снова уменьшился в последних поколениях до наших дней.
Но между 570 и 1005 гг. н. э., когда группы цыган покидали Индию, их эффективный размер популяции составлял от 1380 до 6120 человек.
Оценки эндогамии
Цыгане демонстрируют более высокие показатели эндогамии, чем европейцы или северные индийцы, даже невзирая на то, что они сильно смешаны с европейцами.

Степень генетической однородности в геномах цыган сопоставима с популяциями Южной Индии, которые, как известно, следуют культурным традициям по заключению браков между близкими родственниками. Вероятно, что и у цыган браки заключались преимущественно в пределах определённой социальной группы. Также у цыган отчетливо прослеживается эффект основателя или снижение и смещение генетического разнообразия, а также отмечены последующие, так называемые узкие места в виде сокращения генофонда, которые цыгане переживали в своей демографической истории. Единственными двумя группами, которые демонстрируют более высокие показатели эндогамии и инбридинга, чем у цыган, были южноиндийские популяции ирула и веллалар, говорящие на дравидийских языках. А между группами европейских цыган, балканские цыган показали меньшую степень гомозиготности, чем остальные группы цыган. Это может быть объяснено большей численностью цыган в местах, через которые они попали в Европу, а последующие мигранты, испытывали снижение численности популяции после отделения от балканских цыган. А теперь перейдём к последствиям эндогамии.
Наследственные заболевания у цыган
Высокий уровень эндогамии, присутствующий в геноме цыган, возможно, увеличил вероятность переноса вредных мутаций и аллелей. Проще говоря, увеличились риски наследственных заболеваний. Из 48 известных вариантов мутаций, связанных с такими заболеваниями, 11 было обнаружено только у 40 добровольцев из последней работы. При этом 9 вариантов имели европейское происхождение, указывая на основную европейскую составляющую, а два на южноазиатскую. Среди южноазиатских, один вариант rs121918355, связан с первичной врожденной глаукомой. А второй вариант rs104894396, ассоциирован с множеством заболеваний, связанных с потерей слуха и глухотой.
Хочу обратить внимание на этот раздел комментаторов, которые всеми средствами защищают эндогамию в своём обществе, под предлогом: "зато мы знаем своих предков". Посмотрите какой ценой может обойтись иллюзия знания.

Ведь как показывает практика, даже при высоком уровне эндогамии, от генетического состава образующей популяции, может остаться только 20%... И вряд ли представители этих популяций знают кем были эти 80%, Но также нужно понимать, что у разных популяций и условия развития отличаются...
Предпочтения в смешении
Как уже упоминалось, у европейских цыган обнаружено два источника предков: западно-евразийский, включающий европейские, ближневосточные и кавказские популяции, а также южноазиатский, близкий к пенджабцам. При сравнении пропорций предков в Х-хромосомах и аутосомах можно определить какого пола представители вносили больше своего вклада в цыган. По результатам выяснилось, что мужчинам Ближнего Востока и Кавказа нецыганского происхождения, очень приглянулись цыганки. Да и в целом больший вклад нецыганского происхождения у цыган принадлежит мужчинам. Хотя в меньшей степени к цыганам присоединялись и женщины из мест, где они проходили. А среди европейских популяций, наиболее полюбивших цыганок, оказались мужчины Пиренейского полуострова, Прибалтики и Восточной Европы.
Заключение
Демографическая история цыган характеризуется серией узких мест, до 44% от первоначальной численности популяции и примесей до 80%, которые начали происходить, когда прото-цыгане покинули Индию, и продолжались после их прибытия на Балканы, а позже, и в результате расселения по Европе. Примечательно, что сильная примесь у цыган не стерла признаков того, что цыгане испытали сильное сокращение генофонда в начале расселения. Однако эта западноевразийская примесь частично компенсировала предыдущую утрату генетического разнообразия цыган.
Изучение генетического профиля цыган в мировом контексте ставит их между южными азиатами и европейцами, причём ближе к европейцам, что подтверждается и предыдущими данными о примесях. А предки цыган происходят из региона Пенджаб в северо-западной части Индийского субконтинента, несмотря на обнаруженную неоднородность населения в этом регионе, что согласуется и с лингвистическими данными. По разным оценкам, прото-цыгане отделились от населения, говорящего на пенджаби от 2 до 1,5 тыс. лет назад, и позже покинули Индию, через несколько поколений, когда генетически уже стали отличаться от пенджабцев.
Интересно, что несмотря на то, что цыгане считаются изолированной популяцией с частыми случаями сокращения генофонда, расщеплениями и эндогамией, они всё же являются сильно смешанной популяцией со второй родиной на Балканском полуострове, если говорить упрощённо.

Источники:
1. Erica Bianco, Guillaume Laval, Neus Font-Porterias, Carla García-Fernandez, Begoña Dobon, Rubén Sabido-Vera, Emilija Sukarova Stefanovska, Vaidutis Kučinskas, Halyna Makukh, Horolma Pamjav, Lluis Quintana-Murci, Mihai G Netea, Jaume Bertranpetit, Francesc Calafell, David Comas, Recent common origin, reduced population size, and marked admixture have shaped European Roma genomes, Molecular Biology and Evolution, , msaa156, doi.org/10.1093/molbev/msaa156
2. Font-Porterias N, Arauna LR, Poveda A, Bianco E, Rebato E, Prata MJ, et al. (2019) European Roma groups show complex West Eurasian admixture footprints and a common South Asian genetic origin. PLoS Genet 15(9): e1008417. doi.org/10.1371/journal.pgen.1008417
В новой работе палеогенетиков под руководством Сванте Паабо, из германского Института эволюционной антропологии Общества Макса Планка в Лейпциге, исследователи сообщают о третьем по счёту неандертальском геноме, секвенированном с высоким охватом.
Объектом исследования стала представительница неандертальцев из Чагырской пещеры в южной части Алтайского края, проживавшая там около 60-80 тыс. лет назад.

Неандертальцы, населявшие Европу и Западную Азию, распространились на восток, до Алтая на юге Сибири, но географическое происхождение и время прибытия популяций неандертальцев остаются нерешенными. В результате раскопок в Чагырской пещере было обнаружено 90 тыс. каменных и костяных артефактов и 74 фрагмента окаменелостей неандертальца, а также останки животных и растений, извлеченные из слоёв, возрастом от 59 до 49 тыс. лет. В предыдущей работе уже описывались находки из пещеры, но об этом упомяну чуть позже.

А по результатам новой работы, представительница Чагырской пещеры оказалась генетически ближе к европейским неандертальцам, чем к более древним неандертальцам из Денисовой пещеры, расположенной в 106 км восточнее Чагырской. Помимо этого, в Денисовой пещере были найдены останки девочки, у которой отцом был денисовец, а мать неандерталка, как раз относительно тесно связанная с более поздним неандертальцем из хорватской пещеры Виндия. Так что эти древние популяции встречались в Алтайском крае. А связь с Виндией указывает на замену неандертальских популяций. Молекулярно-генетическое датирование показало возраст образца Чагырская 8 в пределах около 80 тыс. лет. Но вот оптически стимулируемое люминесцентное датирование археологического слоя находки показало возраст в 60 тыс. лет. Это может указывать на то, что генетические даты, основанные на текущей частоте мутаций у современных людей, неверны. Возможные объяснения могут быть связаны с тем, что у неандертальцев частота мутаций была ниже, или она изменилась у современных людей в последнее время.
Отношение к другим неандертальцам и денисовцам
Чагырская 8 имеет больше производных аллелей с неандертальцем из хорватской пещеры Виндия под номером 33.19 и другими более поздними неандертальцами Европы и Кавказа чем с более древним неандертальцем Денисова 5 Алтая. При этом представительница Чагырской пещеры, разделяет меньшее количество производных аллелей с другими неандертальцами Европы, которые там жили примерно в то же время, что и хорватский неандерталец.
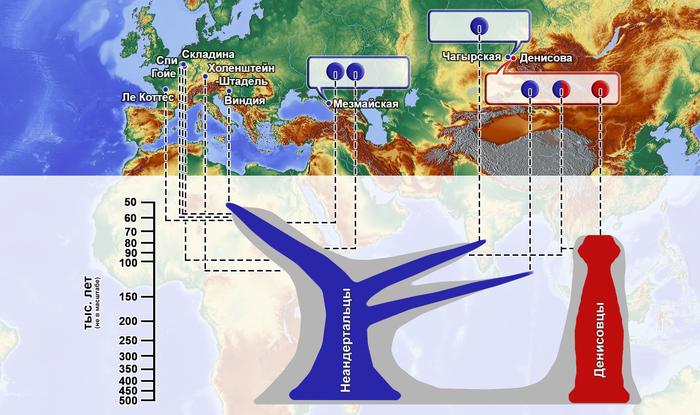
Также Чагырская 8 разделяет больше аллелей с дочкой денисовца и неандерталки из Денисовой пещеры, что в совокупности со схожим распределением производных аллелей с денисовцем-Денисова 3, как и у Виндии 33.19, указывает на более близкую связь Чагырской неандерталки с матерью девочки из Денисовой пещеры.
Подтверждение данных археологии
В февральской работе этого года, российские археологи, совместно с учёными из Германии, Польши и Канады отмечали, что инструментальная сборка Чагырской пещеры, хронологически и технологически пересекается с микокской индустрией Европы и Кавказа.
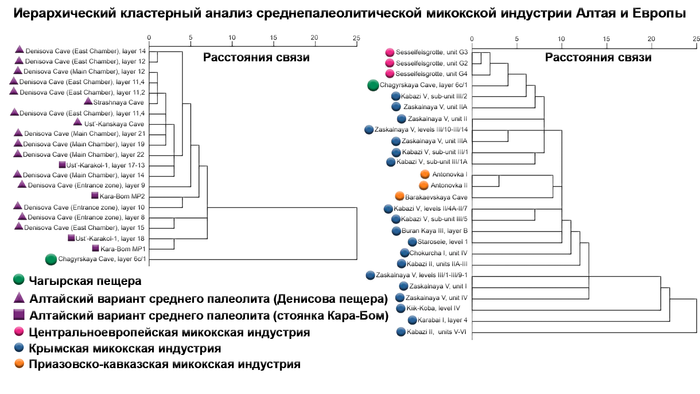
При этом популяции микокской культуры считаются специализированными охотниками на лошадей и бизонов, адаптированными к окружающей среде степей и предгорий. Авторы предыдущей работы связывали их присутствие на Алтае с миграцией неандертальцев из восточной Европы на восток вдоль евразийского степного пояса в холодных и засушливых условиях около 120-80 тыс. лет назад. А отсутствие микокских артефактов в Денисовой пещере в отложениях от 59 до 49 тыс. лет, объяснялось тем, что Денисова и Чагырская пещеры в разное время были заняты двумя отличающимися неандертальскими популяциями. Эти предположения, основанные на материальной культуре, теперь подтверждены и генетическими данными.
Отношения с современными людьми
Учитывая, что современные неафриканские популяции имеют около 2% неандертальской ДНК, в результате примеси между 50 и 90 тыс. лет назад, Чагырская 8 разделяет больше аллелей с современными популяциями людей за пределами Африки, чем более древний образец Денисова 5, но меньше чем Виндия 33.19.
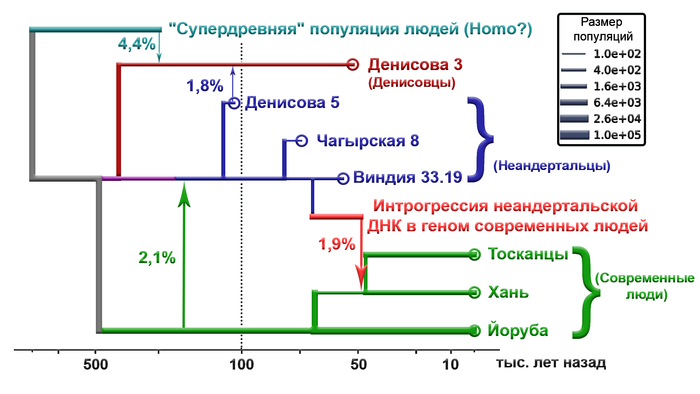
А это указывает на то, что неандерталец из Хорватии ближе к неандертальским популяциям, которые внесли большую часть ДНК в популяции современных людей.
Небольшой размер популяции и инбридинг
По гомозиготным участкам определённой длинны, исследователи определили, что алтайские неандертальцы жили в подгруппах, численностью 60 и менее участников.
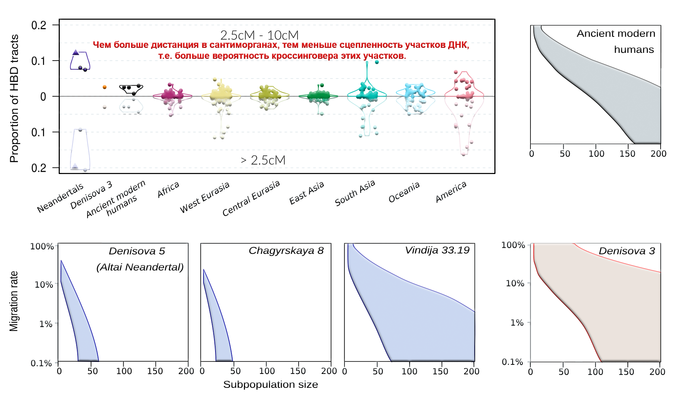
При этом денисовцы и более поздние группы современных людей Алтая были более многочисленны, примерно от 100 человек и более. Помимо этого, хорватские неандертальцы, по-видимому, также жили в группах большей численности, но это различие является лишь незначительно статистически значимым.
Генетические особенности у неандертальцев
Используя три неандертальских генома с высоким охватом, исследователи идентифицировали 993 фиксированные несинонимичные замены среди 889 генов.
При анализе генов, преимущественно экспрессируемых в разных областях мозга, исследователи обнаружили, что гены, экспрессируемые в полосатом теле мозга у подростков в возрасте от 12 до 19 лет, могут быть связаны с ослабленным отрицательным отбором (это когда удаляются вредные аллели). Однако некоторые из белков, кодируемых этими генами, могли быть мишенями для позитивного отбора. При этом такие замены возможно были ликвидированы отрицательным отбором у людей.
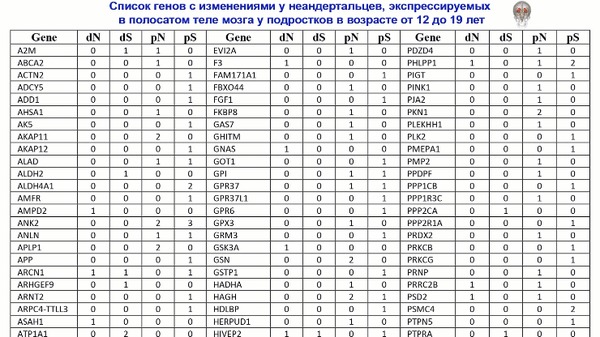
Помимо полосатого тела, гены, экспрессируемые в различных частях теменной, префронтальной и в первичной соматосенсорной коре подростков-неандертальцев, несут более фиксированные замены в своих регуляторных областях, чем гены, экспрессируемые в других областях мозга и в другом возрасте. При этом некоторые гены теменной коры связаны с речью и математическими способностями.
Также гены, вовлечённые в положительный отбор у неандертальцев, могут быть связаны с развитием нервной системы, иммунитетом и заживлением ран, а также функций митохондрий. Некоторые подобные области генома, также подвергались положительному отбору и у современных людей.
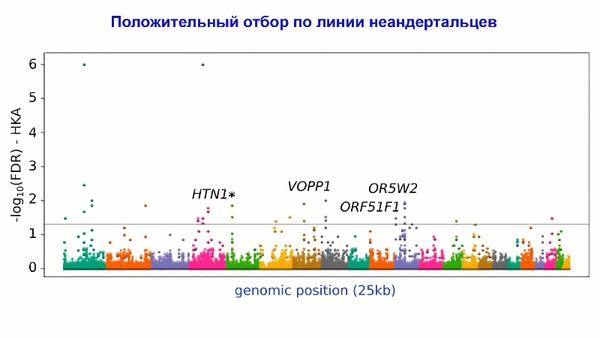
Кроме того, среди главных фенотипов, связанных с изменениями у неандертальцев, есть аномалии в частях скелета, где подчёркивается морфология неандертальцев, как к примеру переносица и грудная клетка.
Выводы
Представительница неандертальцев из Чагырской пещеры, оказалась более тесно связана с хорватским неандертальцем Виндии и другими, более поздними неандертальцами Западной Евразии, чем с более древним неандертальцем, представленным образцом 5 из Денисовой пещеры. А стало быть Чагырская 8 связана с неандертальцами, которые мигрировали на восток где-то между 120 и 80 тыс. лет назад и принесли свою материальную культуру. Что подтверждается и артефактами, обнаруженными в Чагырской пещере. При этом эта новая волна неандертальцев на периферии своего ареала, сталкивалась с популяциями денисовцев, проживавших в Сибири. Не обошлось и без дружбы организмами. На что указывает их совместное потомство в виде девочки Денисова 11.

Хотя может эти контакты были и не совсем добровольными, мы-то имеем лишь результат, а по своей воле или нет, неизвестно... Но это не удивительно учитывая малочисленность и изолированность популяций алтайских неандертальцев.
Помимо этого, побочным эффектом малочисленной популяции, было наличие генов экспрессируемых в полосатом теле мозга, которые могли нести специфичные для неандертальцев изменения, но были невыгодны современным людям. По мере получения более качественных неандертальских геномов, появится и возможность всесторонне изучить гены и группы генов, которые несут функционально значимые изменения для неандертальцев, а также сравнить их с современными людьми.
Источники:
1. A high-coverage Neandertal genome from Chagyrskaya CaveSvante Pääbo, Fabrizio Mafessoni, Steffi Grote, Cesare de Filippo, Viviane Slon, Kseniya A. Kolobova, Bence Viola, Sergey V. Markin, Manjusha Chintalapati, Stephane Peyrégne, Laurits Skov, Pontus Skoglund, Andrey I. Krivoshapkin, Anatoly P. Derevianko, Matthias Meyer, Janet Kelso, Benjamin Peter, Kay PrüferProceedings of the National Academy of Sciences Jun 2020, 117 (26) 15132-15136; doi.org/10.1073/pnas.2004944117
2. Archaeological evidence for two separate dispersals of Neanderthals into southern SiberiaAnatoly P. Derevianko, Kseniya A. Kolobova, Richard G. Roberts, Victor P. Chabai, Zenobia Jacobs, Maciej T. Krajcarz, Alena V. Shalagina, Andrey I. Krivoshapkin, Bo Li, Thorsten Uthmeier, Sergey V. Markin, Mike W. Morley, Kieran O’Gorman, Natalia A. Rudaya, Sahra Talamo, Bence Viola, Proceedings of the National Academy of Sciences Feb 2020, 117 (6) 2879-2885; doi.org/10.1073/pnas.1918047117