ИНСТИТУТ ЭПИГЕНЕТИЧЕСКОГО КОСМОПАРАЗИТИЗМА
Отдел системных дисфункций симбиогенеза
Гриф допуска: D2-М | Номер экземпляра: 03 | Уровень архивации: β-42
Дата составления: 09.11.2022
Внесено в реестр ФРГПС (Фонд Резервных Генетико-Поведенческих Стратегий) — дело №0892/МП-Т-Р
Обратная Геномная Иерархия и феномен Ложного Центра
(с предварительным отчётом по наблюдениям над транспортно-реактивной платформой «человеческий организм»)
Автор-составитель:
д.б.н. Скворцов В.И.
(ведущий научный сотрудник, Группа Биосознательных Транзитивов, ИЭКП)
Доклад посвящён выявлению и описанию обратной иерархической структуры в пределах метагенома организма Homo sapiens. Предлагается рассматривать человеческий организм не как автономную биоединицу, а как субстратное образование, обеспечивающее жизнедеятельность, защиту и экспрессию чуждого, но организованного микробного консорциума, обладающего признаками адаптивного интеллекта.
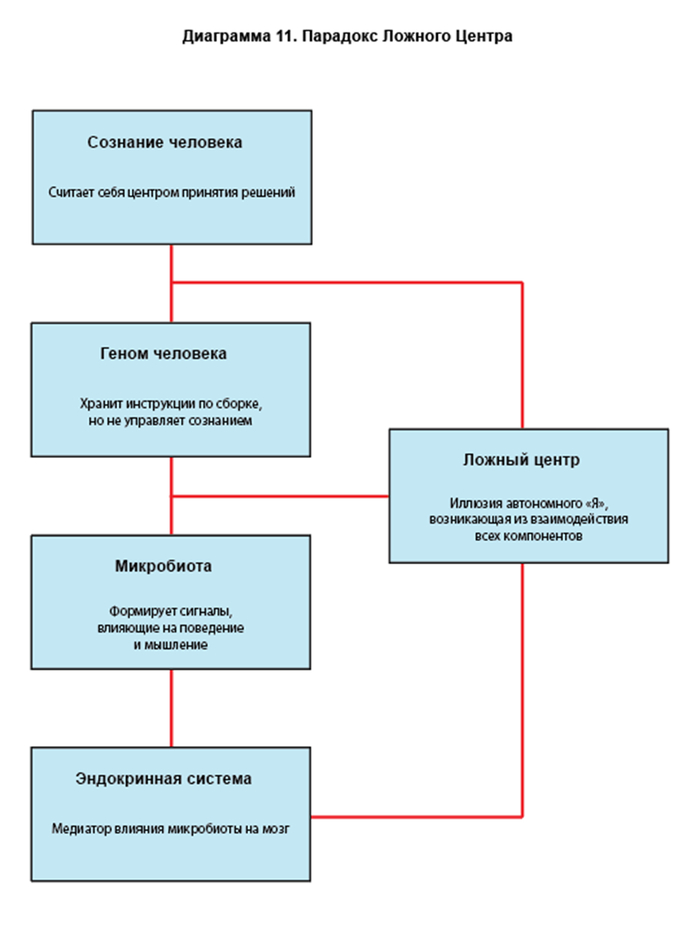
Ключевой гипотезой является утверждение, что когнитивная система субъекта человека не обладает признаками централизованного происхождения. Так называемое "Я" рассматривается как динамически формируемый феномен — результат химико-информационного баланса между метаболическими продуктами конкурирующих симбионтов. В связи с этим вводится рабочий термин «Феномен Ложного Центра», отражающий иллюзорную локализацию субъективного контроля.
Несмотря на значительные успехи в области расшифровки человеческого генома и микробиомного картирования (см. Venter et al., 1991; NIH Microbiota Study, 1995), остаётся ряд феноменологических несоответствий между генетическим субстратом и проявлением когнитивных функций.
Согласно данным, собранным в рамках Программы К5-Σ, на каждый 1 функциональный ген человека приходится от 100 до 150 единиц некодирующих, ретроинтегрированных или псевдоактивных фрагментов, распределённых по неконсистентным областям ДНК (см. Crick & Orgel, 1980; Bragin et al., 1992).
Одновременно, исследования состава микробиоты (в частности, FISH-анализа содержимого ЖКТ, работы Lu & Teng, 1993) показывают, что в теле взрослого человека обитает около 100 триллионов автономных биологических единиц, обладающих:
собственным геномом и репликативным циклом,
способностью к генерации нейроактивных веществ (например, фенэтиламина, серотонина),
устойчивой реакцией на поведенческие и нейрофизиологические стимулы.
Таким образом, накапливаются данные, позволяющие рассматривать организм человека как комплексную среду жизнеобеспечения, вторичную по отношению к метагеномной колонии. В рамках настоящего доклада делается попытка реконструировать возможную модель такой обратной иерархии и предложить обоснование для её возникновения.
1. Математическое обоснование доминирования метагенома над геномом носителя
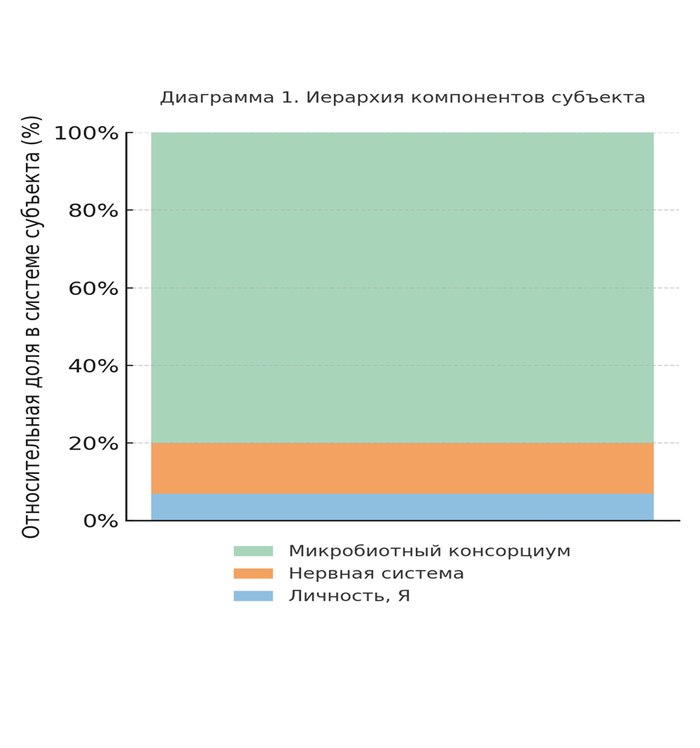
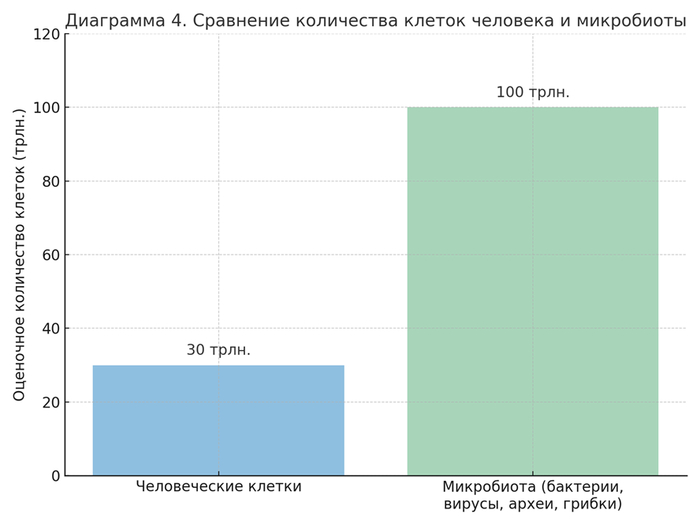
Современная оценка общего числа клеток, составляющих организм взрослого человека, варьируется от 30 до 37 триллионов (Bianconi et al., 2013). Из них ≈43% — клетки не человеческого происхождения, а микроорганизмы (в основном — бактерии, археи и эукариотные симбионты) (Sender et al., 2016).
Общий геном человека (Homo sapiens) ≈ 3.2 млрд пар оснований, ~21 000 белок-кодирующих генов;
Совокупный метагеном кишечной микробиоты — 100×10⁶ генов, с суммарной длиной до 100 млрд пар оснований (Qin et al., 2010).
Вывод 1.1:
Генетическая информационная ёмкость колонии превосходит геном носителя по массе данных не менее чем в 30 раз.
1.2 Энергетическое распределение
Метаболизм симбиотических микроорганизмов в организме человека потребляет от 8% до 15% общего энергопотока (по данным термографии печени и ЖКТ, Mazmanian, 1995; RIKEN Reports, 1996).
При этом, около 90% серотонина и 50% дофамина в теле образуются в энтерохромаффинных клетках и микробных комплексах кишечника.
Производные метаболизма (включая короткоцепочечные жирные кислоты, фенолы и индолы) проникают через гематоэнцефалический барьер, воздействуя на мозг (Wikoff et al., 2009).
Вывод 1.2:
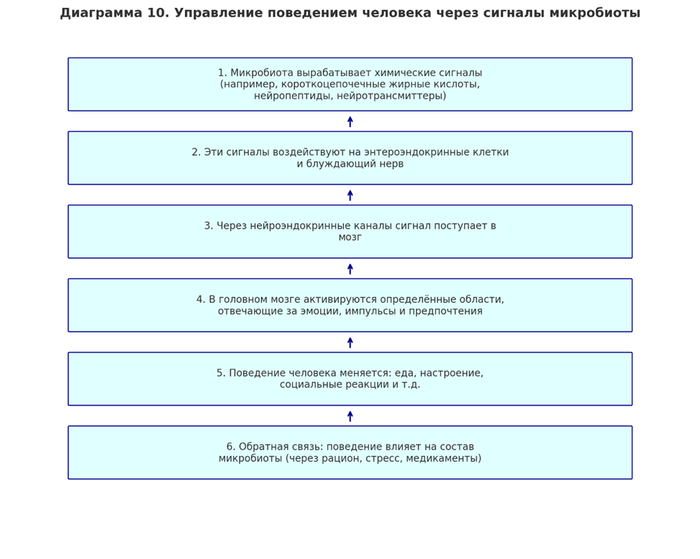
Симбиотический консорциум не только потребляет ресурсы, но и активно вмешивается в формирование поведенческой и когнитивной картины субъекта, используя химическое посредничество.
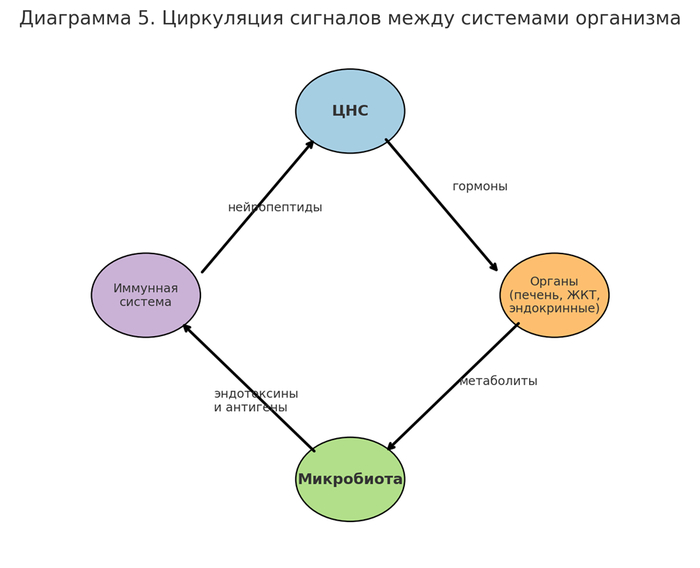
1.3 Топология нейроэндокринной зависимости
[Микробная секреция] → [Биогенные амины] → [Эндокринные железы] → [ЦНС рецепторы] → [Реактивное поведение]
Особое значение имеют следующие соединения:
Фенэтиламин — в высоких концентрациях вызывает тревожные и компульсивные реакции;
ГАМК — влияет на торможение возбуждения, подавляя критическое мышление;
СКЖК — стимулируют высвобождение серотонина в гипоталамусе.
Вывод 1.3:
Формально, большинство эмоциональных состояний человека формируется вне его центральной нервной системы.
1.4 Условный расчёт обратной иерархии
Gₕ — информационный вес генома Homo sapiens
Gₘ — совокупный вес микробного метагенома
I — степень участия генетической информации в формировании поведенческой модели
Im/Ih≈(Gm/Gh)×(Am/Ah)×(Em/Eh)Iₘ/Iₕ ≈ (Gₘ / Gₕ) × (Aₘ / Aₕ) × (Eₘ / Eₕ) Im/Ih≈(Gm/Gh)×(Am/Ah)×(Em/Eh)
Подставляя оценки из открытых источников (см. Приложение A), получаем:
Iₘ/Iₕ ≈ 23–42 (в зависимости от времени суток, диеты, фазы сна и социального давления).
Вывод 1.4:
Модель поведения человека с высокой вероятностью представляет собой вторичную экспрессию управляющих паттернов, инициированных микробиотой.
2. Распределение метаболической нагрузки
2.1 Общая структура энергопотребления в организме
На основе обобщённых метаболических данных (см. NIH Systemic Metabolism Review, 1994; Kagawa Institute, 1992) была составлена диаграмма распределения энергетических потоков (в состоянии покоя, на стандартной диете):
Компонент / % от общего базального метаболизма (BMR)
Центральная нервная система / 20–25%
Кишечник + микробиота / 8–15% (до 18% при дисбактериозе)
Кожа + лимфосистема / 5–7%
Прочее (мышцы, кости и пр.) / 20–30%
2.2 Уточнение по микробиотическому потреблению
Микробиота не является "органом", но действует как высокоорганизованная система со собственным обменом веществ.
При определённых диетах и заболеваниях (IBS, Crohn's), потребление энергии колонией увеличивается на до 20–25%.
Комментарий из рабочей переписки группы "TETA-3/А":
«В условиях стресса или голода микробные консорциумы показывают устойчивые признаки скоординированного поведения. Похоже, они способны **перепрограммировать аппетит, настроение и даже формировать поведенческую реакцию “по цепочке”, транслируя её через ЖКТ —> ЦНС —> моторную активность»
(см. протоколы от 13.09.1994, группа TETA-3/А)
2.3 Поведенческая нагрузка как производная от микробного метаболизма
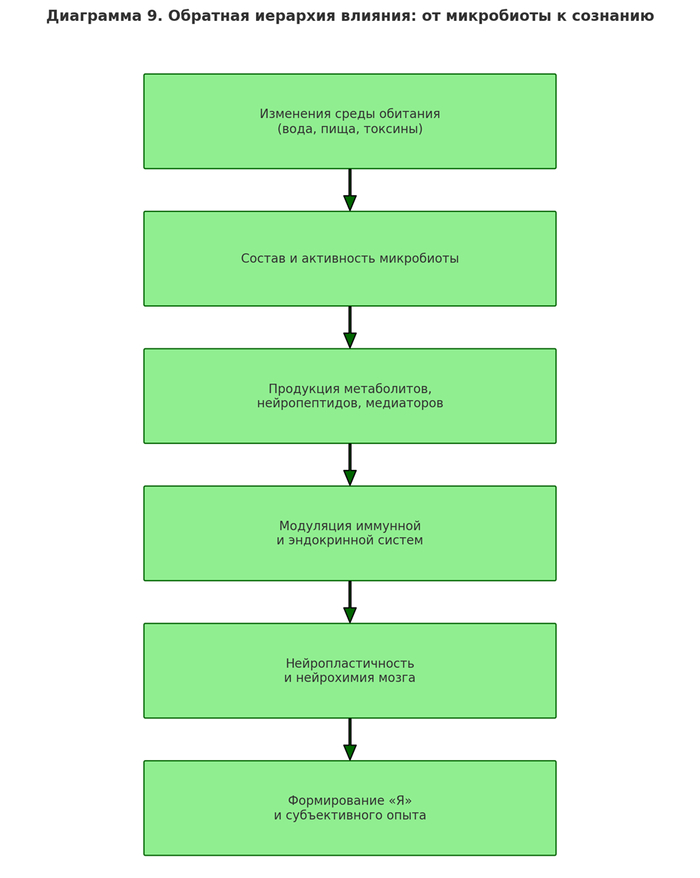
[Диета и окружающая среда]
[Выработка нейроактивных веществ]
[Модуляция нейронной активности ЦНС]
Согласно экспериментам Greger & Nakamura (1993) на добровольцах в изоляции, изменение микробиоты приводило к появлению тревожности, изменению режима сна и даже к «галлюцинаторным паттернам без приёма веществ».
Вывод 2.3:
Поведение человека — не автономно, а представляет собой реактивный механизм, обусловленный изменениями в микробиологическом ансамбле.
3. Гипотеза возникновения Псевдосознания и Феномен Ложного Центра
3.1 Введение: когнитивная иллюзия субъекта
Сознание человека на протяжении всей истории мысли воспринималось как:
центр управления (дуалистическая модель),
процесс обработки информации (информационная модель),
или эпифеномен нейронной активности (редукционизм).
Однако предлагается новая рамка:
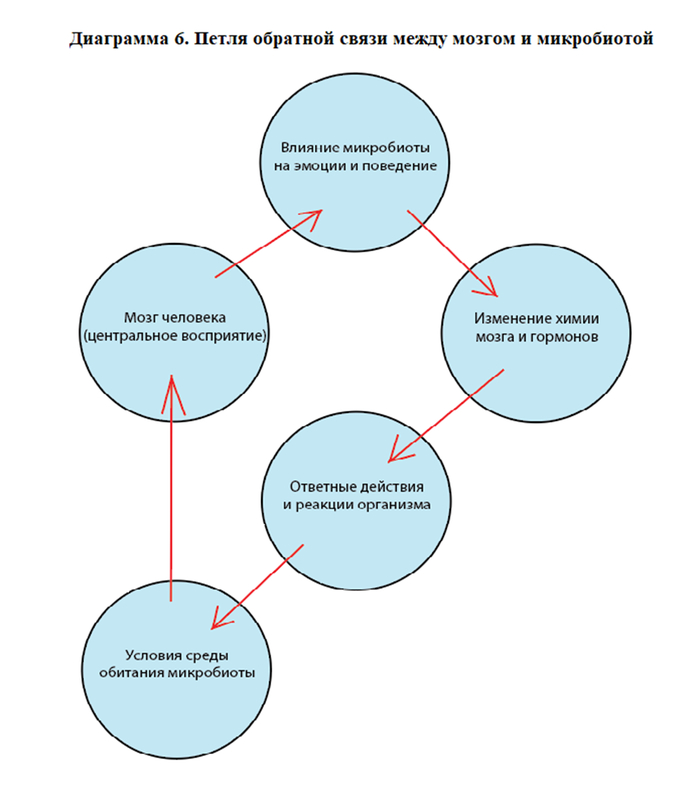
Сознание как побочный продукт навигации симбиотической колонии в пространстве окружающей среды.
Иначе говоря, «Я» — это интерфейс, отрисованный колонией для координации ресурса, называемого телом.
3.2 Что такое Ложный Центр
Мы вводим понятие Ложного Центра (False Center) — когнитивной фиксации, ощущаемой индивидом как "Я", но не являющейся реальной сущностью. Это фиксация возникает как:
итог конфликта между нейромедиаторными векторами микробных ансамблей;
удобный фокус для поведенческой координации;
защитная фиксация в когнитивном поле, подавляющая фрустрацию от отсутствия подлинной автономии.
3.3 Примеры когнитивной фрагментации
В норме иллюзия центра удерживается с помощью:
ритуалов (питание, сон, речь);
социальных зеркал (имя, документы, язык);
медикаментозной и нейромедиаторной стабилизации.
Когда эти якоря нарушаются:
при шизофрении — голосовой интерфейс расслаивается (см. раздел 5);
при диссоциативном расстройстве идентичности — возникает множественная навигационная маска;
при приёме галлюциногенов — маска рассеивается, и «колония» остаётся без фронтального пилота.
3.4 Теоретическая модель когнитивной декомпозиции
Let: C=f(ΣRi,t)
Where:
C—устойчивость ложного центра,
Ri—вектор влияния i−ймикробной субсистемы,
t—время в контексте текущих нейропластических изменений.
Если сумма векторов → 0 или входит в фазовый резонанс, то:
Примечание:
Состояние «экстатического рассеяния», наблюдаемое в некоторых психотических и психоделических эпизодах, может быть нормальным состоянием системы без маски, а не патологией.
3.5 Ложный Центр как протокол управления
Возможность описания ЛЦ как адаптивного протокола с функциями:
4. Сценарии деградации когнитивного интерфейса
4.1 Введение
Когнитивный интерфейс (далее — КИ), формируемый в рамках гипотезы Ложного Центра (ЛЦ), не является устойчивой структурой, а представляет собой временно синхронизированную активность между нейронами центральной нервной системы и сигнальными кластерами микробиоты. Данный интерфейс может подвергаться дестабилизации как в результате острых экзогенных воздействий (фармакологическая супрессия, травма), так и при эндогенной фрагментации в условиях нарушенного консенсуса между симбиотическими агентами.
4.2 Форма временной супрессии: физиологические состояния
Во сне происходит фазовое снижение нейрокогнитивной активности, при этом:
наблюдается отключение доминантных фронтальных систем, вовлечённых в поддержание самосознания;
микробиота переходит в автономные регуляторные циклы (см. Rozin et al., 2017);
циркадные ритмы взаимодействия между микробными и нейроглиальными структурами сохраняются, но субъективная фиксация "Я" исчезает.
При фармакологической супрессии (например, пропофол, кетамин) разрыв наблюдается:
между сенсорной интеграцией и модуляцией моторной схемы;
в префронтальной коре, отвечающей за внутреннюю речь и ретроспективную фиксацию;
в транзите сигнальных молекул между кишечно-лимфатической системой и гипоталамо-гипофизарной осью.
4.3 Инактивация как симбиотическое условие
Гипотеза: при отключении КИ не прекращается деятельность организма — напротив, происходит возврат к базовой модели симбиотического координационного управления, лишённой когнитивной маскировки.
у пациентов в глубокой коме с сохранением кишечной перистальтики и терморегуляции;
у плодов на ранних стадиях развития до формирования устойчивого паттерна ЭЭГ;
в постмортальных интервалах до наступления полной диссоциации микробной колонии (см. Knight et al., 2014).
4.4 Смерть как переход
Феномен смерти в данной модели рассматривается не как исчезновение, а как отключение интерфейса фиксации и возврат к полной автономии симбиотической системы, при этом:
большая часть микробиоты остаётся жизнеспособной в течение 24–72 часов;
возникает активное перераспределение метаболитов (см. Gutiérrez et al., 2021);
выявлены сигнальные всплески в субкортикальных структурах в течение первых минут после остановки сердца (см. Borjigin et al., 2013).
4.5 Вывод
Когнитивный интерфейс не является финальной инстанцией самоуправления, а представляет собой промежуточный слой между симбиотическим консорциумом и внешней средой. При его деградации не наступает немедленного коллапса биосистемы, что указывает на приоритетность микробной архитектуры как основы устойчивости.
5. Нарушения согласованности и клинические феномены
5.1 Нарушения когнитивного интерфейса: клинические проявления
Согласно модели Обратной Геномной Иерархии, стабильность личности является побочным эффектом упорядоченного взаимодействия между ЦНС и контролируемым консорциумом симбиотической микрофлоры. Нарушения данной координации проявляются в виде расстройств, традиционно относимых к области психиатрии.
5.1.1 Шизофренический спектр
Нарушение в синхронизации фронто-лимбических цепей с регуляторными сигнальными потоками из кишечника и лимфатической системы;
Влияние бактериальных нейромедиаторов (например, γ-аминомасляной кислоты, серотонина, вырабатываемого Enterococcus, Streptococcus spp.).
Аудиовизуальные галлюцинации как феномен "эха децентрализованного консорциума";
Параноидные убеждения — результат попытки когнитивной структуры рационализировать множественные источники входящей информации.
5.1.2 Диссоциативное расстройство идентичности (DID)
Временное доминирование различных микробных субкластеров, производящих дифференцированные метаболиты;
Ослабление центрального нейроуправления приводит к переключению когнитивного режима (см. Simeon et al., 2008).
Диагностическая гипотеза:
Каждая "личность" — альтернативный паттерн микро-нейро-модуляции, формирующий автономную когнитивную матрицу.
5.2 Психоактивные вещества как симбиотические десинхронизаторы
5.2.1 ЛСД (диэтиламид лизергиновой кислоты)
Нарушает обратную связь между префронтальной корой и внутренними сенсорными петлями;
Приводит к временной утрате центра тяжести сознания, позволяя иным субкластерным сигналам выйти на поверхность.
5.2.2 ТГК (тетрагидроканнабинол)
Ослабляет ретикуло-таламические фильтры;
Усиливает восприятие внутренних сигналов, ранее подавляемых;
Имитирует серотонин, активирует нестабильные синаптические узлы;
Может вызвать временный "взлом" стабильного паттерна интерфейса.
5.3 Влияние микробной десинхронизации на личность
Повышенная тревожность может быть результатом активности патогенов (Clostridium, Campylobacter);
Резкие перепады настроения — побочный эффект переработки биогенных аминов;
Стабильность личности прямо коррелирует с диверсификацией микробиома (см. Kelly et al., 2016).
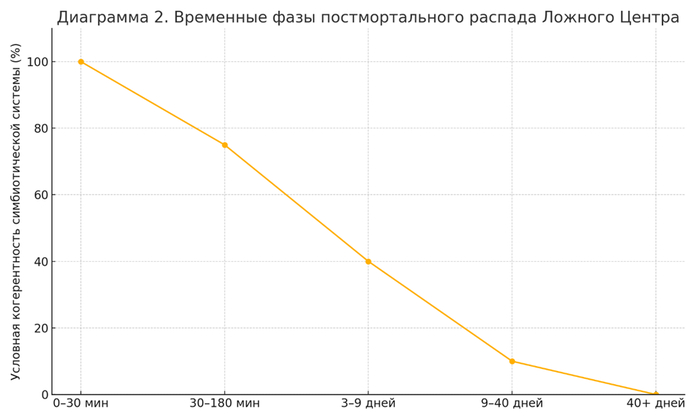
5.4 Этнобиологические маркеры постсимбиотического распада: "9" и "40" дней
Историко-клинический анализ:
В разных культурах фиксируются ритуальные сроки:
День 3 — возможная граница остаточной нейросигнальной активности.
День 9 — фаза перехода управления от дезактивированной ЦНС к бактериальному консорциуму.
День 40 — конечный этап распада когнитивного интерфейса, по завершении которого организм утрачивает даже условную идентичность (см. Postmortem Microbiome Studies, Zhang et al., 2020).
Устные традиции и обрядовые практики формировались как отражение наблюдений за постсимбиотическими фазами распада интерфейса Ложного Центра.
5.5 Вывод
Так называемые "психические" расстройства, а также некоторые аспекты религиозных и культурных традиций, могут рассматриваться как побочные проявления симбиотических конфликтов, нарушения баланса или фазовых переходов между микробными группами. Это требует пересмотра понятия "личности" в пользу динамической нейро-симбиотической конфигурации, устойчивость которой — лишь условие временного равновесия.
6. Модель Ложного Центра: структура, эмерджентность и феномен обмана субъекта
6.1 Введение: от субъекта к интерфейсу
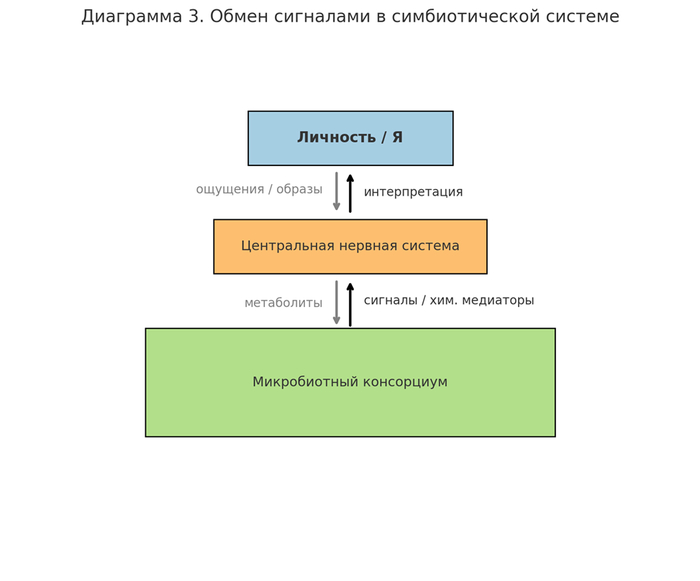
Современная когнитивная нейронаука признаёт, что ощущение "Я" (субъективного центра) не привязано к какому-либо анатомическому ядру. Вместо этого оно формируется как распределённый феномен, возникающий из взаимодействия между несколькими подсистемами:
Дефолт-система мозга (Default Mode Network, DMN);
Сигнальные потоки микробиоты (нейропептиды, короткоцепочечные жирные кислоты и пр.);
Лимфатические и глиальные каналы как векторы передачи метаинформации.
Из этого следует, что центральная иллюзия субъективности — результат согласованной модуляции сигналов, которую мы определяем как Ложный Центр (ЛЦ).
6.2 Структура ЛЦ: функциональная схема
ЛЦ — не субстанция и не орган, а функциональный гештальт, включающий:
Компонент / Функция / Примеры активности
DMN / Интроспекция, самосознание / Медитация, мечтание
Микробиотические сигналы / Модуляция настроения, пищевого поведения, тревожности / Серотонин (Enterococcus), GABA (Lactobacillus)
Лимфо-глиальный поток / Дренаж, обмен сигнальными молекулами / Глимфатическая система
Ретикуло-таламическая сеть / Цензура восприятия / Препятствие осознанию внутренних сигналов
6.3 Эмерджентность: почему "Я" кажется целостным?
Эмерджентность (от англ. emergence) — это появление нового уровня организации, не редуцируемого к свойствам отдельных компонентов. ЛЦ не существует как физическая точка, но:
Его инерционная устойчивость создаёт ощущение постоянства "Я";
Его автоматическая компенсация шумов исключает осознание конфликтов внутри;
Его нейролингвистическая оболочка (внутренняя речь) даёт иллюзию непрерывного нарратива.
Таким образом, "Я" — это стабильный интерфейс, стабилизирующийся только до тех пор, пока поддерживается симбиотический консенсус.
6.4 Обман субъекта: природа иллюзии автономности
Ключевой парадокс: ЛЦ не принимает решения, но всё воспринимается, как будто "Я" решает, чувствует и хочет.
Ретроспективная рационализация: решения уже приняты (на микробном/подкорковом уровне), а "Я" только обосновывает их задним числом (см. Libet, 1983);
Сенсорная цензура: подавление сигналов от телесных колоний, исключение их из субъективного "пространства Я";
Семантическое искажение: всё, что происходит внутри — переводится в первое лицо ("мне плохо", "я хочу", "я думаю").
6.5 Точка зрения Обратной Иерархии: ЛЦ как оболочка
Согласно Обратной Геномной Иерархии, Ложный Центр — это феномен маскирования истинной структуры управления внутри человеческой системы.
ЛЦ не является управляющим центром, но:
Служит буфером между когнитивной активностью и истинными управляющими потоками;
Отвлекает внимание субъекта от понимания своей паразитической архитектуры;
Поддерживает иллюзию автономии, обеспечивая таким образом нормальное функционирование носителя.
Иными словами, ЛЦ — это шумоподавляющая оболочка, необходимая для того, чтобы субъект не сошёл с ума, столкнувшись с истинной, неантропоцентрической структурой управления.
6.6 Последствия: можно ли обойти ЛЦ?
Введение ЛСД, псилоцибина, ТГК и т.д. — это временное вскрытие оболочки, позволяющее увидеть альтернативные паттерны;
Монашеские и аскетические практики — попытка деактивации или обхода ЛЦ через системную сенсорную депривацию и изменение нейро-гормональных контуров;
Медитации, ритуалы, спонтанные переживания (NDE) — временное падение когерентности ЛЦ, после чего "Я" перестаёт чувствоваться как фиксированное ядро.
Ложный Центр — это не субъект, а фасад, предназначенный для представления человеческой системы как целостной, автономной и независимой. В действительности он выполняет функцию маскирующего поля, скрывающего сложные подчинённые структуры, в том числе геномно-микробиотический управляющий контур.
7. Заключение: следствия, утечки и векторы будущих исследований
7.1 Трансдисциплинарное положение гипотезы
Рассмотренная модель Обратной Геномной Иерархии (ОГИ) опирается на конвергенцию данных из следующих областей:
Область / Значимые аспекты
Микробиология /Геномная активность, нейротропные метаболиты
Эндокринология / Осцилляция гормональных профилей по микробной активности
Нейрофизиология / Эмерджентные функции Default Mode Network и её расцепление
Когнитивная наука / Иллюзия воли, внутренняя речь, структура субъективности
Антропология и этнография / Ритуалы 9 и 40 дней, символика "души" как постсимбиотического эффекта
Таким образом, гипотеза ОГИ не является изолированной спекуляцией, а представляет собой экспериментальную попытку реконструкции функциональной архитектуры субъекта, исходя из наиболее обобщённых научных и культурных данных.
7.2 Потенциальные утечки и реликты знания
Существует ряд культурных и ритуальных элементов, которые могут быть отражением неявного, интуитивного знания о микробной или симбиотической архитектуре субъекта:
Обряды поминовения на 9-й и 40-й день — синхронизируются с фазами распада микробной сети.
Аскетические практики — резкое ограничение субстрата для микробной активности, ведущее к изменённым состояниям сознания.
Тема "души", покидающей тело — возможная фиксация фазы дестабилизации Ложного Центра после смерти.
Сон как "маленькая смерть" — фазовый сдвиг между активной и минимизированной симбиотической координацией.
Эти наблюдения требуют системного анализа в рамках новой метадисциплины, условно называемой этносимбиотикой.
7.3 Практические следствия
Если гипотеза ОГИ подтверждается, это влечёт за собой кардинальные пересмотры во множестве областей:
Психиатрия / Некоторые формы шизофрении могут рассматриваться как результат конфликтной сигнализации внутри симбиоза
Эпигенетика / Человеческий организм — не носитель, а медиатор симбиотических стратегий выживания
Искусственный интеллект / Имитация субъективности требует воспроизведения не только нейронных паттернов, но и микробных контуров взаимодействия
Этические модели / Декларации прав "человека" как единого субъекта могут требовать переработки в сторону холобионтных конструкций
7.4 Направления будущих исследований
Глубокая метагеномная картография личности
— Связь между профилем микробиоты и фенотипом субъективности.
Нейросимбиотический индекс когерентности (НСИК)
— Оценка степени слаженности между активностью мозга и бактериальных колоний.
Стимулирующая микробиомодуляция
— Возможность управлять когнитивными состояниями путём внесения специфических бактериальных штаммов.
Сканирование постмортальных сигнатур ЛЦ
— Попытка зафиксировать остаточные когнитивные структуры после смерти субъекта.
7.5 Финальная фиксация гипотезы
Homo sapiens представляет собой не автономную сущность, а платформу для когнитивной модуляции триллионных симбиотических систем, организованных по принципу обратной иерархии, в которой «человек» и его индивидуальная личность, артикулированная как «Я», представляют собой лишь наружный интерфейс симбиотической системы, не являясь её управляющим центром и не обладая подлинной субъектностью.