Места нахождения останков: Африка (Лангебаанвег, Эфиопия, Танзания и долина реки Омо).
Цивета Лики является самым крупным известным на данный момент представителем семейства Виверровых. Она достигала размера маленького леопарда, имея при этом рост 59 см и вес около 40 кг.
Внешне напоминает современные азиатские виды. Считается, что этот вид тесно связан с видом Civettictis civetta (африканская цивета).
Микростоникс был крупнее любой современной дикой свиньи (его масса доходила до 300 кг, а длина черепа до 50 см). По строению он в целом напоминал современных представителей семейства свиней, но имел и характерные отличия от них. Самое заметное отличие — у микростоникса были совсем небольшие клыки. Вместо развитых клыков альвеолярные гребни над верхними клыками (особенно у самцов) были очень крупным и выглядели, как пара бугров на морде. Видимо, они были всеядными.
Предполагается, что микростониксы населяли берега рек и переувлажнённые леса.
Микростоникс появился в позднем миоцене в Азии. Он вскоре стал обычным представителем гиппарионовой фауны и за несколько миллионов лет расселился по Евразии от Испании до Китая.
К началу плиоцена ареал микростоникса начинает понемногу сокращаться. Предполагаемые причины — иссушения климата и смена экологической обстановки. Его место занял род кабаны.
Арандаспиды (лат.Arandaspida) — небольшая группа вымерших панцирных бесчелюстных из класса парноноздрёвых (Pteraspidomorpha), которой присваивают ранг от отряда до подкласса. Название образовано от названия австралийского народааранда и др.-греч. ἀσπίς — «щит» (обычное окончание для названий вымерших бесчелюстных).
По внешнему виду арандаспиды были рыбоподобными животными, но имели только один плавник — хвостовой. Достигали длины 35 см. Были покрыты тонким панцирем. Примечательны парностью отверстия для теменного глаза (хотя эти отверстия имеют и другую интерпретацию).
В составе этой группы различают от 2 до 8 видов. Арандаспид сближают с разнощитковыми, а иногда и включают в их состав.
Чаще всего к арандаспидам относят 4 рода, 2 из которых — предположительно, поскольку они известны только по мелким фрагментам. В каждом из этих родов описан только один вид:
Sacabambaspis janvieri Gagnier et al., 1986, известный из Боливии, Аргентины, Австралии и Омана. Назван в честь типового местонахождения — боливийского города Сакабамба. Жил в конце нижнего ордовика (лланвирнский и карадокский век, около 470 млн лет назад). Известен по останкам намного лучшей сохранности, чем остальные арандаспиды, в том числе по практически целым экзоскелетам.
Andinaspis suarezorum Gagnier, 1991 из ордовика или девона Боливии (недалеко от местонахождения Sacabambaspis). Известен по единственному обломку; принадлежность к арандаспидам не доказана.
Arandaspis prionotolepis Ritchie & Gilbert-Tomlinson, 1977 из центральной Австралии (Алис-Спрингс). Назван в честь живущей в этих местах этнической группы. Жил в начале верхнего ордовика (карадокский век, около 450 млн лет назад). Известен по отпечаткам передней части экзоскелета в мелкозернистом песчанике;
Porophoraspis crenulata Ritchie & Gilbert-Tomlinson, 1977 оттуда же. Кроме того, подобные ему окаменелости известны из раннеордовикских пород (аренигский ярус, около 480 млн лет назад), и это самые древние находки арандаспид). Получил название за большие поры на бугорках панциря. Известен только по мелким фрагментам; принадлежность к арандаспидам не доказана.
Кроме того, в родах Sacabambaspis, Arandaspis и Porophoraspis, вероятно, есть как минимум по одному неописанному виду. Не исключено, что к арандаспидам относится и ещё один боливийский вид — Pirchanchaspis rinconensis Erdtmann et al., 2000, но его систематическое положение очень неясное.
Почти целая окаменелость
Арандаспиды имели вытянутое каплевидное тело длиной до 35 см, покрытое хорошо развитым экзоскелетом. Брюшная сторона была более выпуклой, спинная — более плоской. Единственный плавник — хвостовой. Он хорошо изучен только у Sacabambaspis, у которого имел длинную, но узкую центральную лопасть (куда заходила хорда), более короткую спинную и ещё меньшую брюшную.
Глаза находились на самом конце головы. В них были окостенения — склеротические кольца. Между глазами расположены ноздри, а позади — пара отверстий, которые обычно интерпретируются как пинеальные.
Рот арандаспид был на нижней стороне головы. У них было до 10 пар жаберных мешков, которые открывались наружу отдельными отверстиями, а по другим данным — в общие проходы, открывавшиеся одним отверстием с каждой стороны.
Реконструкция Сакабамбасписа
Эндоскелета у арандаспид не было или почти не было, зато был хорошо развит экзоскелет. Переднюю часть тела покрывали две большие костные пластины (спинная и брюшная), а заднюю — сильно вытянутые вертикальные чешуйки. Длина передних пластин составляет около половины длины животного, а толщина очень маленькая (у Arandaspis < 0,1 мм). Они цельные (без следов слияния отдельных элементов) и не имеют признаков роста. Это означает, что они появлялись уже у взрослого животного.
Между спинной и брюшной пластиной с каждой стороны был ряд из 15—20 небольших многоугольных пластинок, между которыми находились жаберные отверстия. Кроме того, много рядов мелких костных пластинок было с нижней стороны рта. На спинном и брюшном щитке, а также на мелких чешуйках видны маленькие, но хорошо развитые каналы боковой линии.
Все элементы экзоскелета образованы аспидином — бесклеточной костью. В них различаются 3 слоя: нижний (пластинчатый), средний (сотоподобный или сетчатый) и верхний (бугорчатый).
Снаружи панцирь был орнаментирован бугорками. Для родов, известных только по обломкам (Andinaspis и Porophoraspis) их форма — главный отличительный признак. У Arandaspis они вытянутые, сужаются к концам и имеют продольный гребень, а у Sacabambaspis, Andinaspis и Porophoraspis в разной мере напоминают дубовые листья, причём у последнего пронизаны большими порами.
Формой и орнаментацией панциря арандаспиды напоминают разнощитковых и астраспид, а чешуйками хвостовой части — анаспид.
Позади глазниц арандаспид, в передней части спинного щита, расположена пара небольших отверстий. Они есть и у Arandaspis, и у Sacabambaspis. Обычно их интерпретируют как отверстия для органов пинеального комплекса — светочувствительной структуры, из которой у многих древних и некоторых современных позвоночных развивается теменной глаз. Таким образом, у арандаспид таких глаз было два: одно отверстие вмещало пинеальный орган, а другое — парапинеальный. Это очень редкая ситуация среди позвоночных (кроме арандаспид, парное или сдвоенное пинеальное отверстие известно только у некоторых ископаемых рыб — ряда плакодерм, палеонисцид, поролепообразных и ранних двоякодышащих). Кроме того, глазоподобность обоих органов пинеального комплекса сохранилась и у большинства миног (хотя пинеального отверстия у них нет).
По другой интерпретации, эти отверстия у арандаспид представляют собой выходы эндолимфатических протоков, которые есть на панцире и некоторых других остракодерм. В пользу этого говорит то, что они расположены дальше назад, чем обычно для пинеального отверстия.
Арандаспиды жили в мелком море. Поскольку они не имели стабилизирующих плавников, их движение, вероятно, было неуклюжим и напоминало движение головастика. Тем не менее они были самыми прогрессивными известными позвоночными своего времени.
Как и для других остракодерм, для арандаспид предполагается придонный образ жизни и питание детритом и микроорганизмами. На это указывает положение рта снизу головы и (как и у других остракодерм) отсутствие челюстей.
Арандаспиды — древнейшие позвоночные, которые известны по довольно полным скелетам (от более древних находили только мелкие обломки панциря или отпечатки бесскелетного тела). Самые древние фрагменты, вероятно, принадлежащие арандаспидам, имеют возраст около 480 млн лет (начало ордовика), а самые молодые — 440 млн лет (конец ордовика). Причина их вымирания неизвестна; возможно, они исчезли из-за оледенения.
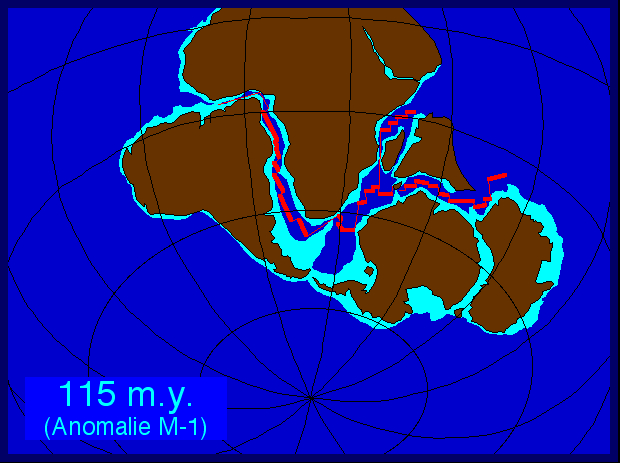
В отличие от других ранних остракодерм (живших в основном в водах Лаврентии), арандаспиды известны только из Гондваны (а именно из прибрежных отложений окружавших её морей). Окаменелости этих животных встречаются в Южной Америке (Боливия и Аргентина), Австралии (центр континента) и на Аравийском полуострове (Оман). Эти места были далеки друг от друга и в ордовике, хотя все находились на краю Гондваны.
Гондвана. Находки арандаспид известны с восточного, западного и северо-восточного края.
Окаменелости арандаспид встречаются редко, но к этой группе относятся почти все ордовикские остракодермы Гондваны. В водах этого континента остракодермы были редки и в последующие времена. Кроме арандаспид, оттуда известны только телодонты (очень широко распространённая группа, возникшая, видимо, в водах Лаврентии и к концу ордовика проникшая в моря Гондваны) и питуриаспиды (маленькая эндемичная для Австралии группа из раннего — среднего девона).
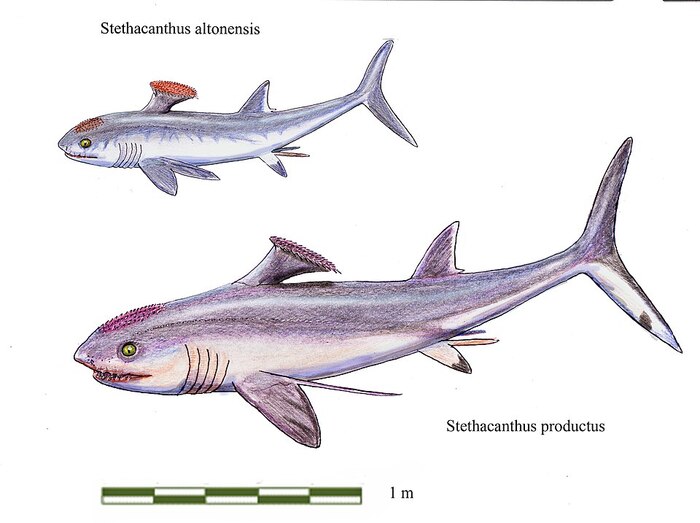
Достигали длины 0,7—2 м. Из-за диковинной формы спинного плавника стетакантов в шутку называют «гладильная доска». Только самцы обладали причудливым спинным плавником, а у самок плавник был обычным, как у современных акул. Назначение этого странного спинного плавника, как и коротких шипов на голове и передних плавниках, неизвестно. Возможно, он играл роль в привлечении самок.
По данным сайта Paleobiology Database, на август 2021 года в род включают 12 вымерших видов:
Stethacanthus altonensis (St. John & Worthen, 1875) [syn. Physonemus altonensis St. John & Worthen, 1875]
Stethacanthus compressus Newberry, 1897
Stethacanthus concavus Ginter, 2018
Stethacanthus depressus (St. John & Worthen, 1875) [syn. Physonemus depressus St. John & Worthen, 1875]
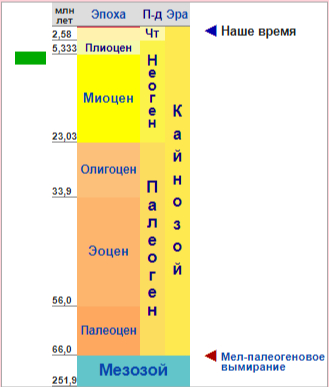
Ихтиозавры являются наиболее адаптированными к водному образу жизни пресмыкающимися. Аналогичной степени приспособленности смогла достичь только одна группа млекопитающих — китообразные. Уникальные характеристики, связанные с водным образом жизни, наблюдаются уже у древнейших известных ихтиозавров из нижнего триаса (шпатский подъярус оленёкского яруса). К таким приспособлениям относятся удлинённая морда, очень большие глаза, преобразованные в плавники конечности и сильно вогнутые позвонки.
Скелет Ихтиозавра
Ключевой эволюционной особенностью ихтиозавров является живорождение, наиболее ранние документальные свидетельства существования которого относятся к анизийскому ярусу среднего триаса. Имели эвриапсидное строение черепа, произошли от диапсидных предков. Зубы сменялись неоднократно в течение жизни. Конечности использовались для поддержания равновесия и контроля направления. У хвоста было две лопасти, нижняя из которых поддерживалась позвоночным столбом. У типичного ихтиозавра были очень большие глаза, защищённые костяным кольцом, говорящим о том, что охотились они ночью. В связи с этим некоторые виды обладали огромными глазами (до 20 см в диаметре). Дополнительно, по-видимому, имелись какие-то кожные рецепторы, подобные боковой линии, на что указывают следы нервов и сосудов на костях черепа. Кожа, лишённая чешуи, для лучшего скольжения в воде была, возможно, покрыта слизью. Хорошо приспособлены к движению с высокой скоростью, как современный тунец. По крайней мере некоторые ихтиозавры предположительно были хорошими глубоководными ныряльщиками, как современные киты. Наиболее вероятная окраска — тёмный верх и светлый низ с синеватым оттенком. Самый крупный вид, описанный по обнаруженным окаменелостям — шонизавр из позднего триаса полярной Канады. В 2003 году описан скелет длиной около 23 метров, но в среднем длина составляла 2—4 метра.
У некоторых ранних ихтиозавров были зубы, с помощью которых они питались моллюсками — аммонитами, наутилоидеями и кальмарами. Очень похоже на то, что они также питались рыбой, а у части более крупных видов были тяжёлые челюсти и зубы, которые показывают, что они питались более мелкими рептилиями.
Анализ ископаемых остатков стеноптеригия (Stenopterygius), найденного в Германии в шахтах Хольцмадена, датированного возрастом 180 млн лет назад и сохранившего небольшие фрагменты мягких тканей, показал, что внутренние слои кожи переходили в изолирующий жировой слой как у теплокровных животных.
Некоторые ихтиозавры были очень крупными. В 2018 в Англии были найдены фрагменты нижней челюсти ихтиозавра, которые указывали на то, что он имел в длину от 20 до 25 метров, его затем описали как вид Ichthyotitan severnensis, а Ophthalmosaurus icenicus весил около тонны[9].
Типовой вид этой группы был описан в 1821 году как Ichthyosaurus communis De la Beche & Conybeare в составе рода ихтиозавров из нового семейства Ichthyosauridae De la Beche & Conybeare, 1821.
Геохронология Ихтиозавра
Ихтиозавры существовали в течение почти всего мезозоя 250—94 млн лет назад, наибольшего расцвета достигли в юрский период, пока в меловой период их не сменили плезиозавры. В мелу количество видов ихтиозавров резко снизилось, лишь один род — платиптеригиусы (Platypterygius) — дожил до начала позднего мела (единственный описанный представитель меловой группы). Предполагается, что глобальное потепление, произошедшее в середине мелового периода, повлекло за собой обеднение кислородом океанических вод, что привело к «аноксидной катастрофе» и вымиранию не приспособившихся к изменениям климата ихтиозавров.